Axons from the lateral geniculate nucleus of the thalamus enter the primary visual cortex via the optic radiation. The primary visual cortex (V1) is Brodmann's area 17 in the human brain, it is located in and around the occipital pole. It is also called the striate cortex, and the calcarine cortex, the first indicating its anatomical appearance and the second indicating its anatomical location.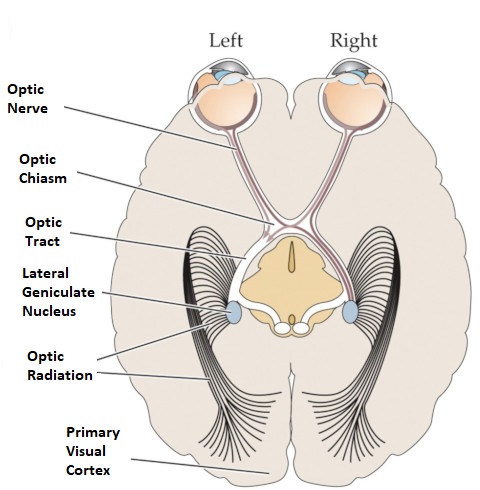  Fibers from the optic radiation enter the primary visual cortex in layer 4, and some K-type axons terminate upstream in layers 1 and 2/3. The axons from each eye that are segregated in the LGN, stay that way in the cerebral cortex, they terminate in different layers and on different neurons. The organization of the early visual processing is schematized in the figure. 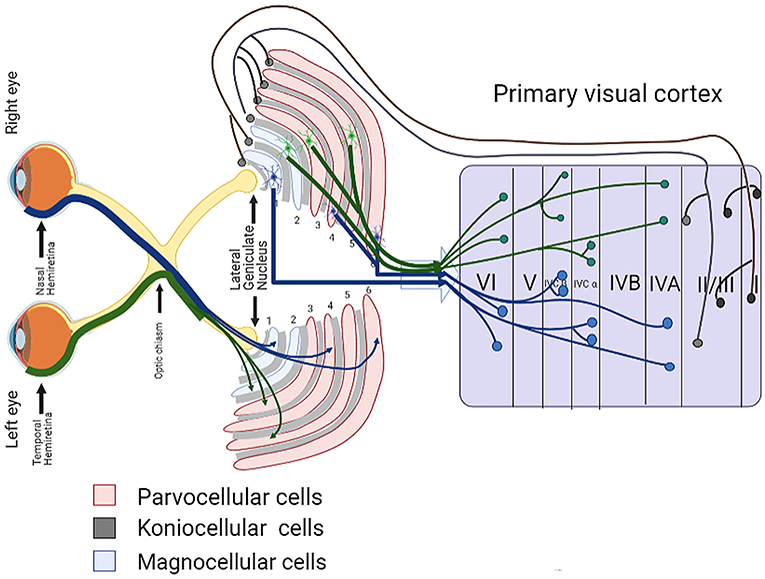 The cerebral cortex in general has 6 layers in humans (with a few exceptions). There are generally two layers of pyramidal cells, labeled layer 5 and layers 2/3. Input from the thalamus arrives in layer 4, and feedback output to the thalamus is sent from layer 6. Other outputs arise from the pyramidal cells and connect widely with many cortical and subcortical structures.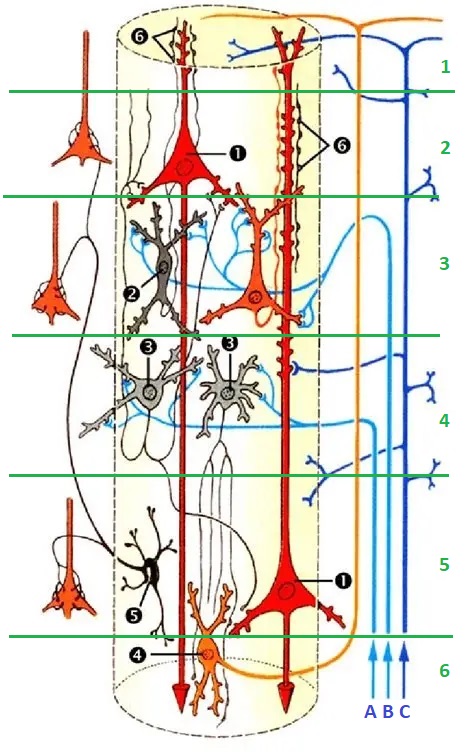 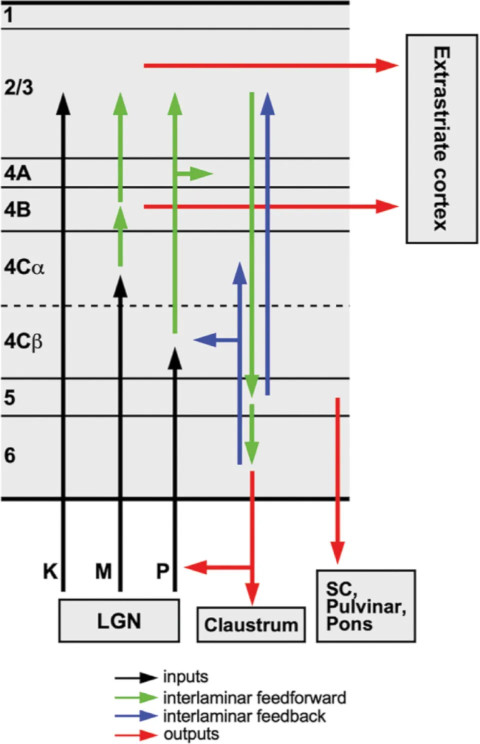 The termination zones of LGN axons in the primary visual cortex are retinotopically organized. The fovea is heavily magnified compared to the periphery. The overall mapping from the visual field to the cerebral cortex resembles a complex logarithmic expansion.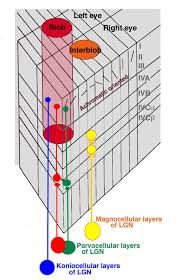 The cortex itself is organized into a matrix of columns, minicolumns, and hypercolumns. In area 17 the axes of this organization are orientation and binocular disparity.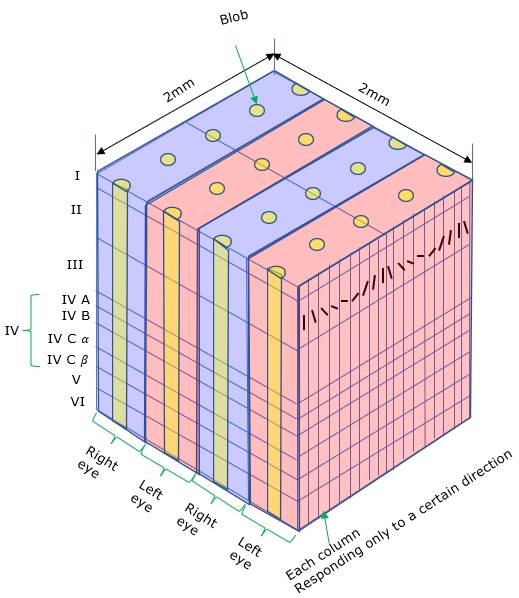 Inserted into this matrix are "blobs" related to color processing, they are regions that stain positive for cytochrome oxidase. Blob cells resemble koniocellular LGN neurons. They are primarily monocular, color sensitive, and characterized by small concentric receptive fields. They cluster within the blobs and receive inputs from kLGN axons. On the other hand, the inter-blob cells are mostly binocular, not color sensitive, and characterized by elongated (rectangular) receptive fields that may or may not have a center-surround organization. The inter-blob cells exhibit ocular dominance and orientation specificity.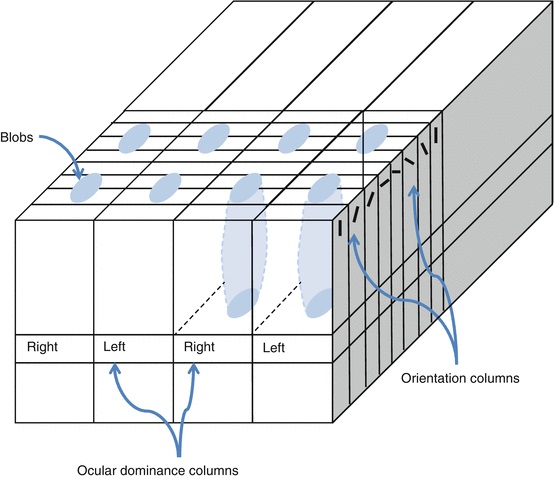 Between blobs, the P pathway from the LGN feeds V1 neurons that are orientation and location specific but not motion sensitive. This pathway primarily processes shape and form. In contrast, the M pathway feeds V1 neurons that respond best to moving stimuli - mostly without a preference for the direction of movement, although some exhibit directional sensitivity.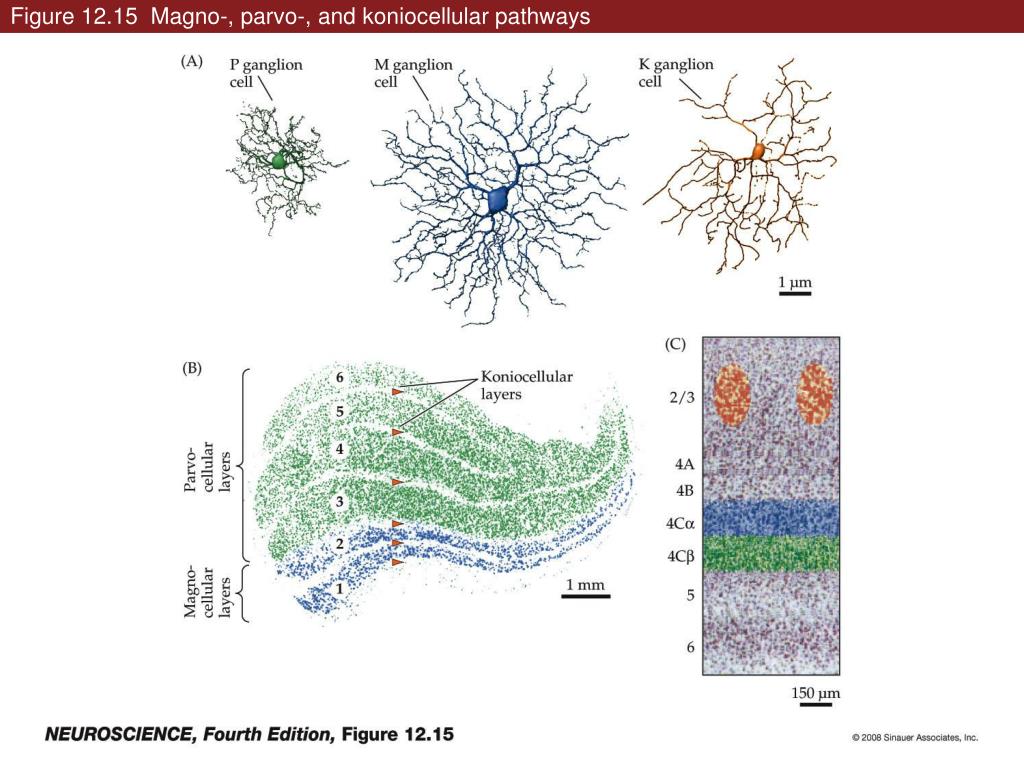 (figure (c) Sinauer Associates Inc, from Neuroscience, 4th edition, Fig 12.15) In real life the matrix of columns in V1 is not as regular as shown above, rather, it is organized into a regular pattern of "pinwheels" as shown in the diagram.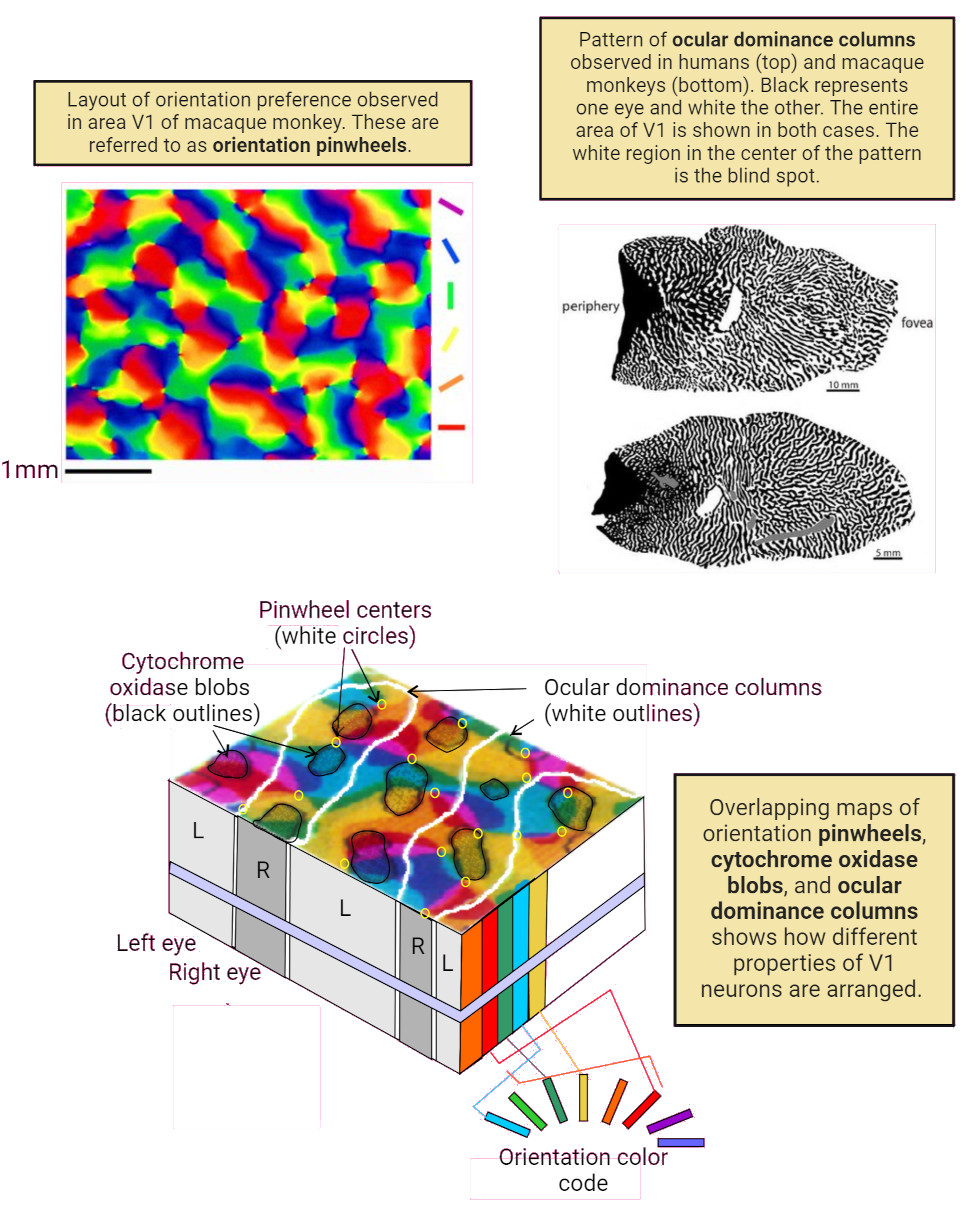 Lesion or removal of area 17 results in total blindness, except for a residual "blind sight" that occasionally perceives vague sensations of motion in the periphery. On the other hand, damage to the extrastriate cortex does not result in a simple loss of vision, rather it results in higher order perceptual deficits including failure to individuate objects and failure to recognize colors and movement. Secondary Visual CortexFrom V1, visual signals travel to V2, area 18 in the Brodmann map of the human cerebral cortex. This area surrounds V1 like a shell, and in fact it is much bigger, it contains more neurons. A schematic of V2 is shown in the figure, and its connection architecture is shown below.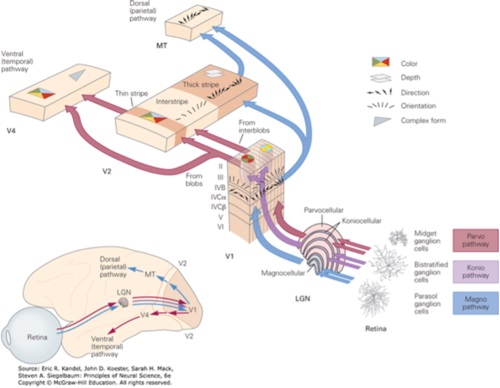 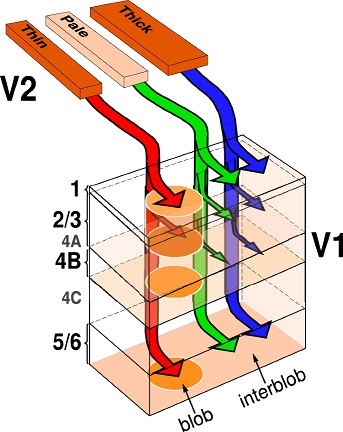 (figure from Federer et al 2021) Higher Visual AreasFrom V2, the visual signal splits into dorsal and ventral pathways. The ventral pathways courses through area MT and enters the parietal lobe. The dorsal pathway connects with V4 and enters the inferior temporal cortex. These pathways and the associated anatomy are shown in the diagrams. (Here Brodmann area 46 is the dorsolateral prefrontal cortex, a large area in the frontal lobe involved in motor planning and cognition).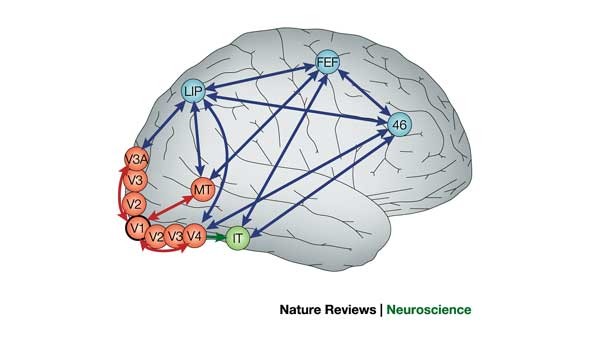 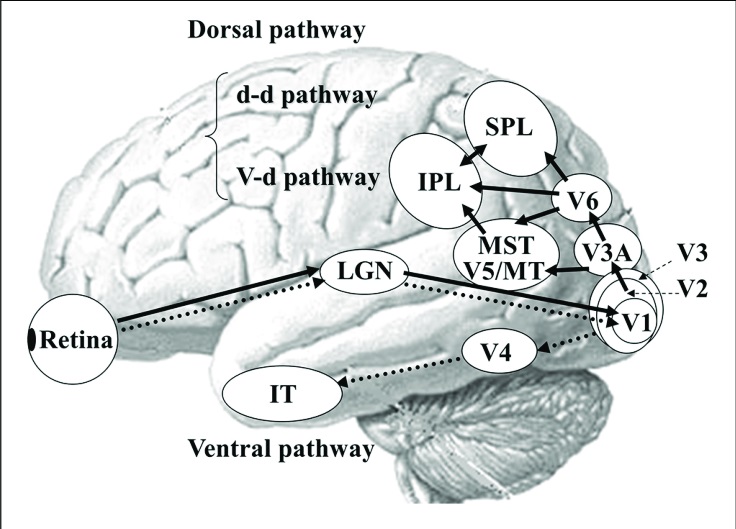 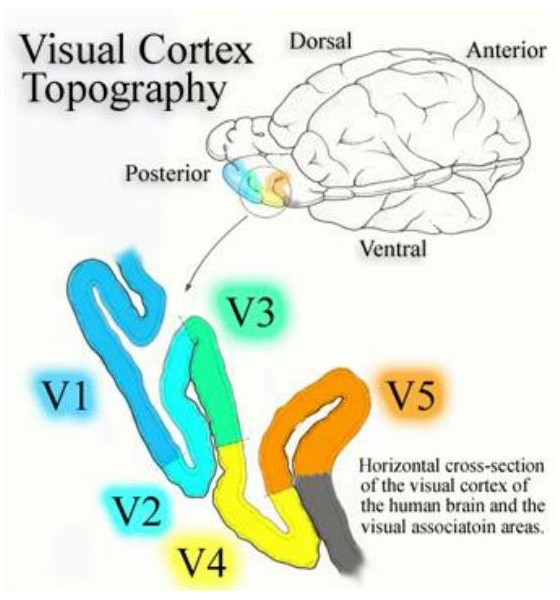 Each of these visual cortical areas is specialized for function. And even within a single area, there are subnetworks specialized for function. Eventually, both visual streams feed into the circuitry around the hippocampus, which reconstructs visual scenes and enters them into short term episodic memory. The entry of visual information into memory is complex and involves more than just scene mapping - like the signal processing itself, it's split up into dedicated pathways in favor of time and accuracy. Visual information from the "what" (ventral) stream is entered differently than information from the "where" (dorsal) stream. Visual IllusionsWe can learn a lot about the visual system by studying the illusions that occur in humans under certain conditions or with certain visual patterns. For example, here are some interesting illusions:   Visual illusions exist at each processing stage along the pathways from the retina through the cerebral cortex. There are some that involve only the retina, like the moon illusion and the retinal shift effect. There are others that involve only the cortex, like these. Is the green dot aligned with the red or is it behind? Are the flashes lined up or are they behind?  Visual illusions like this tell us a lot about visual processing. For example these above reveal the differing conduction delays associated with different aspects of visual information.Next: Visual Streams |