As noted in the previous section, saccadic targeting is organized in the superior colliculus (SC), which connects with the entire range of the peripheral oculomotor system. The SC is a central structure in orienting, which in humans includes body, head, and eye movements. These three movements need to be kept in coordination, and the superior colliculus is an important structure in that regard, as is the cerebellum. However the SC negotiates between different types of eye movements, whereas the cerebellum is both upstream and downstream to this function. In addition to inputs from many areas of the cerebral cortex, the SC receives input from the pars reticulata of the substantia nigra, which in turn is controlled by the caudate nucleus. This latter pathway is important in the disruptions of oculomotor activity related to Parkinson's (Hikosaka et al 2019).
Humans have many strategies available for exploring the environment and targeting stationary or moving objects with eye movements. Saccades and pursuit movements are different outcomes of the same decision process. Some possible interactions of saccades and pursuit are shown in the figure. Typically small saccades allow the eyes to "catch up" with a fast pursuit target. Small errors in target position often generate corrective mini-saccades that are interspersed with smooth tracking.
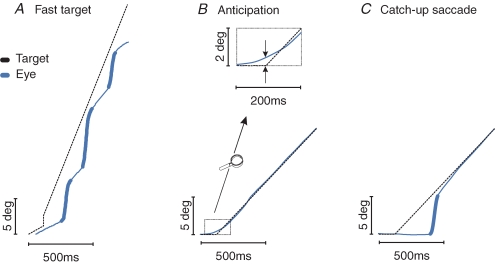
(figure from Orban de Xivry & Lefevre 2007)
Ultimately, targeting of an eye movement is directed by the cerebral cortex, except for some primitive reflexes that mainly involve the level of light. The cortex provides both position and motion information. The position information comes mainly from the frontal and parietal eye fields, while motion information comes from visual area V5. However the SC is a targeting center that coordinates saccades with smooth pursuit movements and instructions from the cerebral cortex as well as predictive information from the cerebellum. The SC is "ahead of" eye movements, it's involved in the selection of future gaze locations, and it reflects covert shifts of attention in the absence of eye movements (Krebs et al 2010). All these roles are important for orienting.
The superior colliculus is a paired layered structure in the roof (tectum) of the midbrain. It is positioned close to both the oculomotor nuclei and the substantia nigra. In birds this area is called "optic tectum", and cross species analysis reveals considerable differences in the mapping of this structure, although the general plan is highly conserved through evolution. Many of the differences are based on differences in the eyes themselves, for example mice have no foveas and cats have streaks. In some organisms (especially those with eyes on the side) the superior colliculus maps the entire visual field, while in humans it only maps the contralateral hemifield. A common element in collicular wiring is the passing of "optic flow" information into the cerebral cortex via the pulvinar of the thalamus. Such flow is created when the organism moves itself in relation to the environment (like, when birds fly, or during approach/avoidance behavior in quadrupeds and humans).
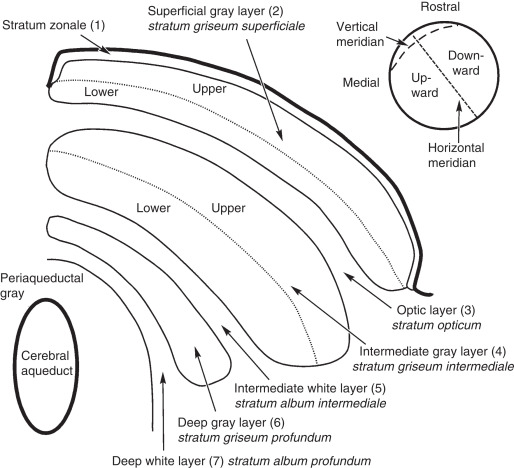
The superior colliculus is generally organized into three major layers, a superficial layer that primarily receives retinotopic input from many parts of the visual system, and intermediate and deep layers that generate a motor map for both visual orienting and other forms of orienting.
The neurons in the superior colliculus are organized vertically in a honeycomb-like columnar arrangement. In the superficial layers, there are narrow-field cells that define the smallest units within this anatomy, and wide-field cells that link neighboring columns. The neurons in the superficial layers then project down into the deeper layers, where they organize a variety of neurons related to eye movement generation and other orienting activities. In addition to the retinotopic map, the superior colliculus maps auditory space and somatosensory space. Sounds localized by auditory means can cause the eyes to move to the corresponding location in visual space, and the sensation of a bug crawling on the skin will invariably cause the eyes to move to that location.
The superior colliculus (SC) is involved with eye movements in many ways. One of its primary tasks is target selection, and in this capacity is has to negotiate between visually driven targets and voluntary targets, and reflex targets like bright spots of light on the retina. In doing so, it must decide what type of eye movement to use to acquire the target. In humans, smooth movements generally occur only within the fixation area, whereas anything more eccentric than a few degrees invokes a saccade. (If it's "very" eccentric the saccade is combined with or replaced by a head movement). The superior colliculus translates position and velocity information from the cortex, into motor vectors for the eye movement system. It does this based on the retinal topography, in other words the relationship between the visual field and the current eye position. Electrical stimulation of a neuron in the superficial layer of the superior colliculus elicits a conjugate contralateral eye movement to the corresponding location in the retinotopic visual field. In other words, the motor map is aligned with the sensory map.
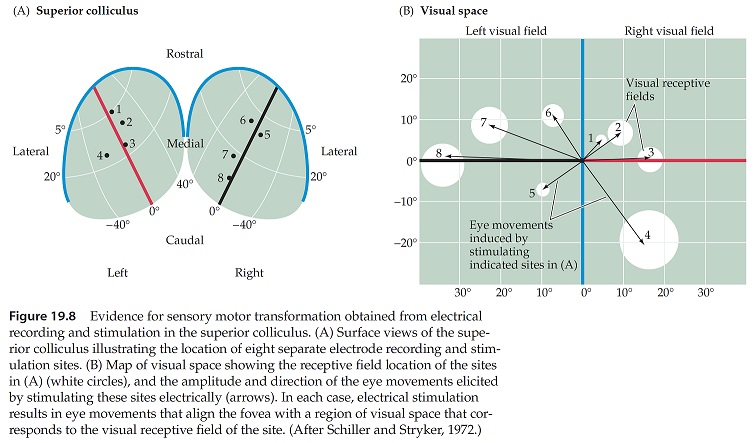
As shown in the figure above, the fovea is mapped into the rostral superior colliculus, while the periphery is in the more caudal area. "Fixation neurons" are found in the rostral area, transitioning to saccade generating neurons in the caudal zone. In between is the decision area for smooth movements, and it is still unclear exactly how this decision occurs. Most likely there is a functional boundary at the fixation border that depends on target velocity.
Generally there are three classes of neurons in the SC:
1. Saccade-related burst neurons that emit a volley of spikes prior to an eye movement,
2. Buildup neurons that ramp activity prior to a saccade, eventually resulting in a burst, and
3. Fixation neurons in the rostral SC that discharge at a tonic rate during fixation and pause during most (but not all) saccades.
In vivo these classes overlap, there is a continuum of neural behaviors spanning these definitions (Gandhi and Katnani 2013).
Saccades are typically very short, less than 100 msec in duration. They typically undershoot the target by about 10%, because they can't be controlled by continuous visual feedback (the delay is too long, the visual information takes about that long to travel through the neural circuits in the cerebral cortex), so control is accomplished by an internal feedback loop based on an efference copy of the commands sent to the motor neurons. At least a portion of this efference copy arises directly from the motor neurons in the abducens nucleus and is passed into the cerebellum by interneurons projecting through the paramedian tract (PMT). In this pathway, the cerebellum contributes to precision targeting by telling the eyes when to stop moving. However there are other connections into the cerebellum directly from the superior colliculus, and from the neural integrators between the SC and the oculomotor neurons. The cerebellum fine-tunes all of these aspects of saccadic movements.
The superior colliculus is involved in all types of eye movements, not just saccades. A subset of SC neurons alter their response characteristics during combined saccade-vergence movements (Walton and Mays 2003), and micro-stimulation of the rostral SC can sometimes induce vergence movements (Chaturvedi and Van Gisbergen 2000) and even lens accommodation (Sawa and Ohtsuka 1994). The activity of neurons in the rostral SC is correlated with smooth pursuit eye movements (Krauzlis 2003), but stimulation there does not evoke smooth pursuit movements. The initiation of smooth pursuit eye movements is till a subject of debate. Centrally, there is a specific area in the frontal lobe called the "frontal pursuit area" that responds to particular pursuit vectors and can be electrically stimulated to induce pursuit movements (Tian and Lynch 1996).
The superior colliculus receives input from both the pursuit and saccadic areas of the frontal eye fields. It is not entirely clear how the "go" signal is achieved, and how exactly the decision is made between a saccadic movement and a pursuit movement. In any case, information from the deeper layers of the superior colliculus is relayed to several pontine nuclei, including the dorsolateral pontine nuclei and the nucleus reticularis tegmenti pontis. Some of the neurons in these areas are velocity and direction sensitive and can be stimulated to change the velocity of pursuit. These pontine neurons project to the vermis and paraflocculus of the cerebellum, in areas distinct from those receiving information about saccades.
While the superficial layers of the SC are retinotopic and process visual information, the deeper layers help coordinate other motor behaviors related to target locations, like approach and avoidance. While the intermediate layers of SC are involved in saccade generation, the deep layers get a body map from the somatosensory cortex. Some of the neurons in the deeper layers of the SC drive head movements as well as body movements, in relation to visually driven and non-visually driven orienting. Head movements take longer and last longer than eye movements, typically SC neuron activity precedes head movement onset by several hundred milliseconds. The SC also has a role in covert attention, that is, allocation in the absence of orienting movements (Krauzlis, Lovejoy, and Zenon 2013).
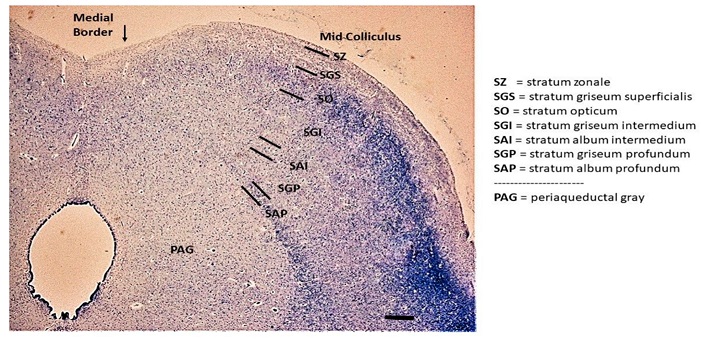
Anatomical OrganizationIt is generally agreed that the superior colliculus has three functional layers, a superficial layer that receives visual input, and two deeper layers (called intermediate and deep) that trigger orienting movements - not just of the eyes, but also of the head and body. Sometimes it is possible to elicit approach or avoidance behaviors by electrically stimulating the superior colliculus. In this figure, the superficial layer consists of the stratum zonale, stratum griseum superficialis, and stratum opticum. The intermediate layer consists of the SGI and SAI, and the deep layer consists of the SGP and SAP.
The superficial layers of the superior colliculus receive direct retinal inputs via collaterals of K-type koniocellular ganglion cells, and collaterals of M-type magnocellular ganglion cells in the periphery, many of which are direction sensitive. However SC neurons in the superficial layers have receptive fields that are considerably larger than those in the retina. In total there are about 100,000 fibers from the retina to the SC, as distinct from over a million from the retina to the LGN.
The superficial layers of the superior colliculus also receive retinotopic inputs from many areas of the visual cortex, including the primary visual cortex (V1, area 17 at the occipital pole), the precuneus (around area 19, V3 in the occipital lobe), and area V5 (MT) in the ventrolateral region of the occipital lobe, just posterior to the junction of the inferior temporal sulcus and the lateral occipital sulcus. In addition to the retinal and cortical inputs into the superficial layers of the SC, there are also inputs from the lateral geniculate nucleus, the pretectal area, and the parabigeminal nucleus.
The superficial SC sends outputs to several areas of the thalamus, including the lateral geniculate nucleus and the posterior and medial nuclei of the inferior pulvinar, as well as the pretectal nuclei and the parabigeminal nucleus. Different groups of neurons in the superficial SC provide these outputs. There is one group that receives input primarily from the visual cortex and projects to the medial subnucleus of the inferior pulvinar (which in turn connects with V5/MT - see Berman and Wurtz 2010). There is another group that receives primarily retinal input and projects to the dorsal lateral geniculate nucleus (May 2006).
There is a direct monosynaptic excitatory projection from the superficial layers to the deeper layers. There is also local GABA-ergic inhibition within the superficial layers, that serves to focus incoming targeting information (the literature sometimes calls this salience detection, since it controls the bursting activity of neurons in the deeper layers).
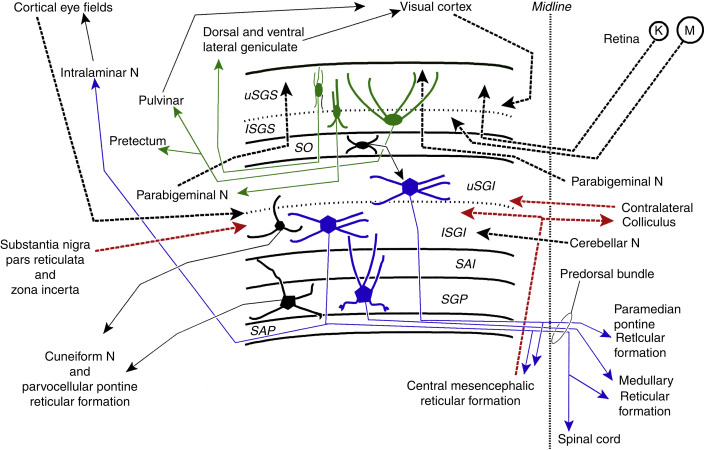
The deeper layers of the SC are also retinotopically organized but have larger receptive fields and don't receive direct projections from the retina. Instead they receive excitatory inputs from almost all over the cerebral cortex (specifically including the frontal eye fields in Brodmann's area 8, the supplementary eye fields in Brodmann's area 6, and the parietal eye fields around the intraparietal sulcus). It is the deeper layers that initiate saccades and other orienting movements. These deeper layers also receive information related to head and body position and motion, from the spinal trigeminal nucleus (conveying information from the head and face), and the contralateral spinal cord and dorsal column nuclei (conveying information about body position). Additionally there is a tonic GABA-ergic inhibitory pathway from the substantia nigra pars reticulata. Other inputs include the hypothalamus, the zona incerta, several areas of the thalamus, the parabrachial nucleus, parabigeminal nucleus (cholinergic), pedunculopontine nucleus (cholinergic), laterodorsal tegmental nucleus (cholinergic), locus coeruleus (noradrenergic), dorsal raphe (serotonergic), and the inferior colliculus. A summary of these connections is shown below.
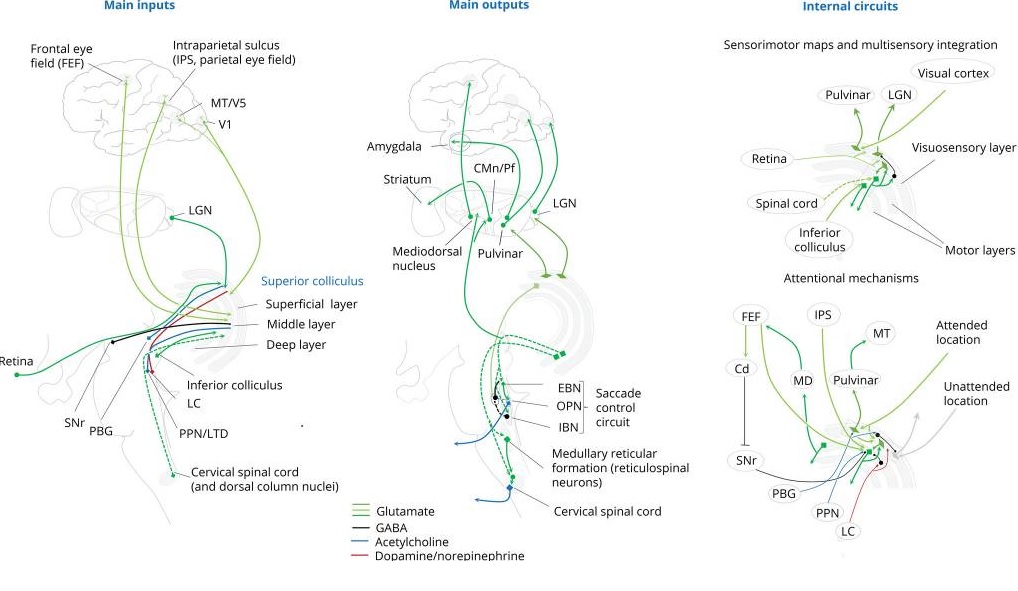
(figure from Benarroch 2023)
The intermediate and deep layers of the superior colliculus control the saccade generating circuitry. They connect contralaterally with the burst and pause neurons in the pons. Collaterals from these fibers then descend to the reticulospinal neurons in the medullary reticular formation (around the gigantocellular nucleus), and the cervical spinal cord. The deeper layers of the SC also send axons rostrally to connect with the pulvinar of the thalamus as well as the lateral geniculate nucleus, the mediodorsal nucleus, and the centromedian and parafascicular nuclei. There are also projections from the deeper SC layers to the substantia nigra pars compacta and the ventral tegmental area, as well as the subthalamic nucleus and the periacqueductal gray.
Internal projections from the deep layers of the SC target the superficial layers. There are direct excitatory connections from the intermediate and deep layers to the superficial layer, which in turn excite the superficial neurons projecting back down to the deeper layers (this is a positive feedback loop - Ghitani et al 2014). Neurons in the deeper layers can also inhibit the superficial neurons projecting to the thalamus (pulvinar and LGN), and this pathway is likely involved in the suppression of retinal motion perception during eye movements (Lee et al 2020).
Neurons within the superior colliculus are organized into a honeycomb arrangement of discrete columns (Illing 1996). In total there are around 100 such columns in rats (Chevalier and Mana 2000), the number in humans is not known.
In addition to the control of the oculomotor system, one of the most important outputs of the superior colliculus is to the inferior nucleus of the pulvinar of the thalamus (posterior and central portions). The pulvinar is involved in visual attention. It receives input from the SC, LGN, and many areas of the visual cortex. In turn it is connected into the circuitry in the dorsal attention stream, as well as the anterior and posterior cingulate cortex and the retrosplenial region. The pulvinar combines target and eye movement information from the SC with visual details from the visual cortex. The figure shows some of the areas of cerebral activation related to the pulvinar.
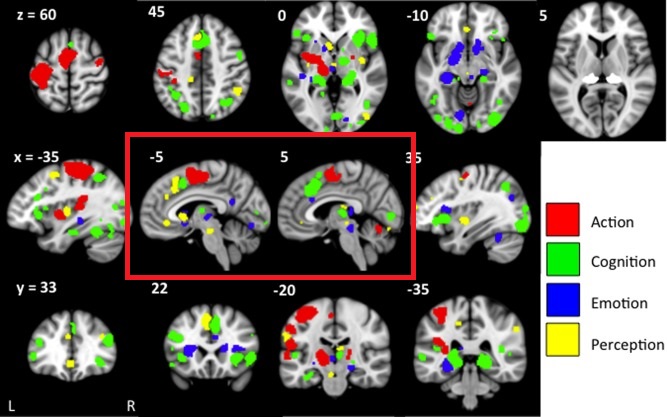
(figure from Barron et al 2015)
A summary of SC wiring and neurochemistry is shown in the figure.
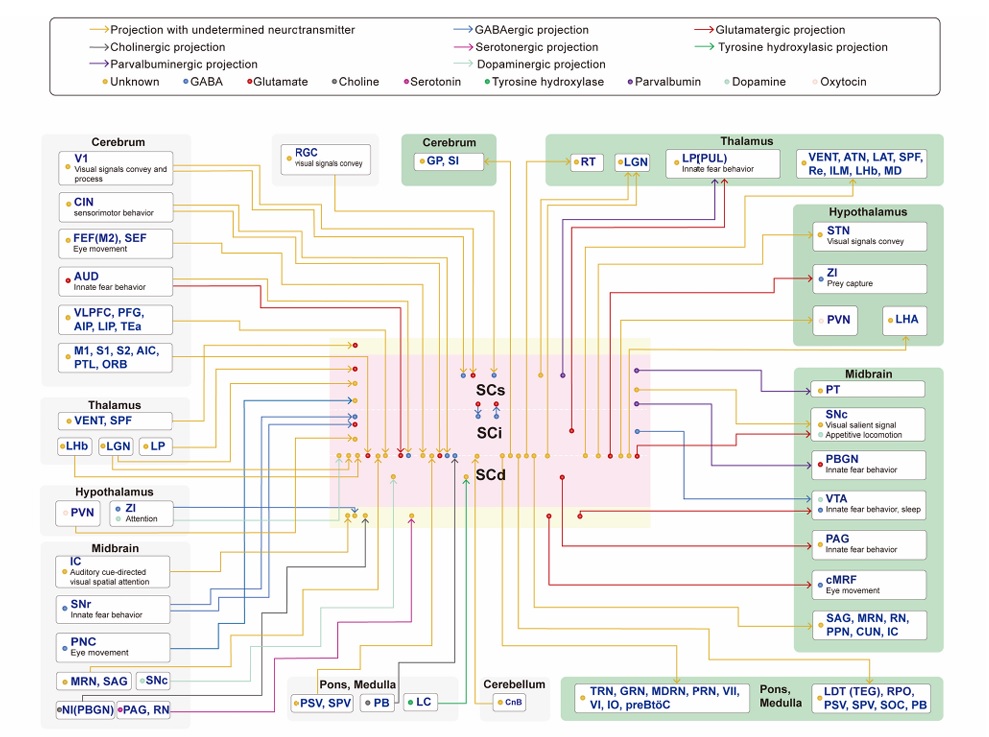
(figure from Liu et al 2021)
Functional OrganizationIn general a visual scene will offer multiple interesting (relevant) targets. The oculomotor system must select between them, and execute exactly one eye movement. The teasing apart of the selection process is somewhat complicated. It can not be done on the basis of neural activity alone, clever experimental paradigms are needed to help us understand what we're seeing.
The evidence indicates that the superior colliculus specifies the location of the target but does not determine the movements that will be used to acquire it. (Krauzlis et al 2004). The superior colliculus can represent multiple possible targets (especially prior to the onset of eye movement), as shown by manipulating target probability during visual search tasks. It must then choose between them prior to releasing an eye movement.
Neurons in the SC generally represent one of three firing patterns: fixation, burst, and build-up. Fixation neurons are active during fixation, and excite the pause neurons in the nucleus raphe interpositus so saccades are suppressed during the fixation period. Build-up neurons increase their firing prior to an eye movement. Build-up neurons represent "candidate targets" for impending eye movements (they do not always generate an eye movement). Burst neurons (usually EBNs) fire just before an eye movement, in a 10-15 msec interval. Once the burst begins the eye movement will almost surely follow. Fixation neurons are usually found in the rostral part of the SC, while the other two types are more prevalent in the caudal portions.
Multiple models have been created to describe the target selection process. The simplest one is "winner take all", where the most relevant stimulus becomes the target. But target selection is a layered process involving low level reflexes in addition to high level proactive choice. There is a direct pathway from the retina to the superficial layers of the superior colliculus, and even after disconnecting other inputs (surgically or chemically) shining a small bright spot of light on the retina can evoke an eye movement to that location.
Anatomically, the rostral pole of the SC contains a map of the contralateral fovea, and it contains fixation neurons, that send a monosynaptic excitatory input to the pause neurons in the NRI. Whereas, the caudal portions of the SC contains neurons that discharge before saccades and project to burst neurons which in turn activate EBN's and IBN's (Shinoda et al 2019). Inhibitory burst neurons also inhibit contralateral pause neurons, providing a disynaptic pathway that contributes to saccade initiation. SC neurons that project to the oculomotor centers send collaterals to the mediodorsal nucleus of the thalamus, which in turn projects to the frontal eye fields. This pathway provides the FEF with feed-forward information about impending eye movements.
In addition to the superior colliculus, there is another midbrain structure involved in target selection and saccade initiation, which is the parabigeminal nucleus. It is a small satellite of the superior colliculus located on the edge of the midbrain at the level of the inferior colliculus (Cui and Malpeli 2003). It is connected with the amygdala as well as the superficial (retinal) portion of the superior colliculus. In rodents it seems to be involved mainly in the initiation of threat avoidance behaviors. Its function in humans is unknown.
Eye Movement InitiationEye movement initiation is a complex process. At one level there is cooperation between the superior colliculus and the cerebellum, but there is also a "traditional motor pathway" involving the caudate nucleus and substantia nigra that engages the cerebellum in a different way, and feeds up into the VPi of the thalamus via the usual somatosensory pathways.
Ultimately it is the cerebral cortex that controls eye movements. At the voluntary level there is involvement of the frontal and supplementary eye fields, and there is another important area near the intraparietal sulcus (area IPS in humans, LIP in monkeys) that engages in visually targeted movements. Furthermore, there is influence from area MT that handles visual movement, and V3 that processes disparity. All of these areas send direct fibers to the superior colliculus from cortical layer 5 pyramidal cells.
At any given time there are multiple visual targets, both those that are present and those that are being attended. When a new target is presented, the brain tends to search it, executing exploratory saccades to different interesting areas in the target. Typically the fixation period is brief, lasting about a third of a second. An example of visual search is shown below.
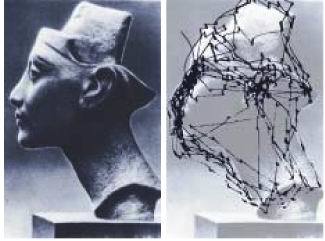
Eye movements are controlled by the attention system. The most salient target is the one the eyes will seek to acquire. Each object in the visual field has an "importance" attached to it (a "value", as it were), and the most important one will win the target selection process. Only a small amount of information can be acquired during each fixation, and in many cases the eyes will revisit the same spot(s) over and over again for additional input.

The frontal and parietal eye fields are part of the dorsal attention network. In addition there is a cingulate eye field in the anterior cingulate cortext, the functions and connections of which are presently unknown in humans.
Eye Movement TerminationMovement termination is also a cooperative process between the superior colliculus and the cerebellum. Burst codes carry velocity and position information but the result is not always exactly accurate, and it is also possible the visual scene has moved before completion of the eye movement. The cerebellum helps guide the eyes to their final target positions. The eyes can either undershoot or overshoot their targets. Both conditions exist in various pathologies, including Parkinson's, MS, ALS, various palsies, and a host of other conditions including stroke. It is instructive to see which pathways are affected in these conditions and how the oculomotor system responds to them. Generally there are disorders in position (hypometria and hypermetria) and disorders related to velocity (slow eye movements).
The superior colliculus is affected by alpha-synuclein and tau neuropathology in Lewy body dementia, shows abnormal responses to visual stimuli in Parkinson's disease, and may contribute to impaired saccades in progressive supreanuclear palsy and the pathophysiology of cervical dystonia (Bennaroch 2023). All of these conditions disrupt the scanning behavior seen in the above two figures. Note that there is a lateral bias in the last figure, the left side is being attended more closely than the right. Such a bias is present naturally in most humans, but can be dramatically exacerbated in pathology. There are commissural connections at all level of the oculomotor system that can be affected by stroke and tumors. Hemispheric neglect is usually contralateral to the affected area. It can occur in the oculomotor system even when the visual system is intact.
Let's take a deeper dive into the cerebellum, as this area works closely together with the superior colliculus and the brainstem oculomotor circuitry.
Next: Cerebellum |