The oculomotor nuclei are organized in the brainstem, at and below the level of the superior colliculus. The general plan is that the abducens and trochlear muscles (lateral rectus and superior oblique, respectively) have their own dedicated nuclei, while the oculomotor nucleus handles the other four muscles. The figure shows the general organization of the oculomotor structures in the brainstem.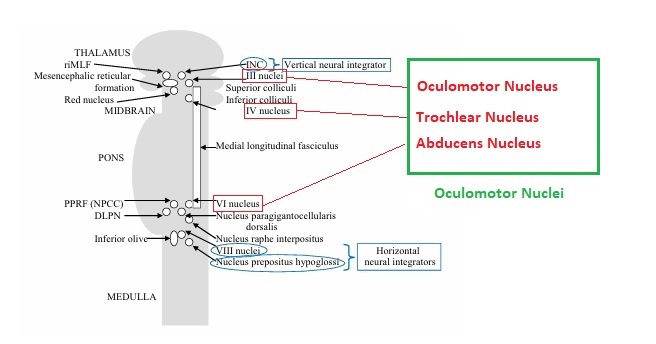 (figure from Sanchez & Rowe 2018) The abducens nucleus controls the lateral rectus muscle, which pulls the eye to the side, away from the midline. This is the most important muscle for horizontal saccadic eye movements. In addition to projecting to the muscles themselves, neurons from the abducens nucleus send axons across the midline to the contralateral oculomotor nucleus, that control the medial rectus muscle in the opposite eye, to keep the eyes in sync. There are also some projections into the floccular region of the cerebellum, and these could perhaps carry a copy of the oculomotor commands for subsequent processing. Motor NeuronsThe motor neurons for the lateral rectus muscle are located in the abducens nucleus (the axons travel in cranial nerve VI, the abducens nerve). Those for the superior oblique muscle are located in the trochlear nucleus (the axons travel in cranial nerve IV, the trochlear nerve). The remaining eye muscles are handled by various subdivisions of the oculomotor nucleus (with axons traveling in cranial nerve III, the oculomotor nerve). The figure shows the subdivisions of the oculomotor nucleus.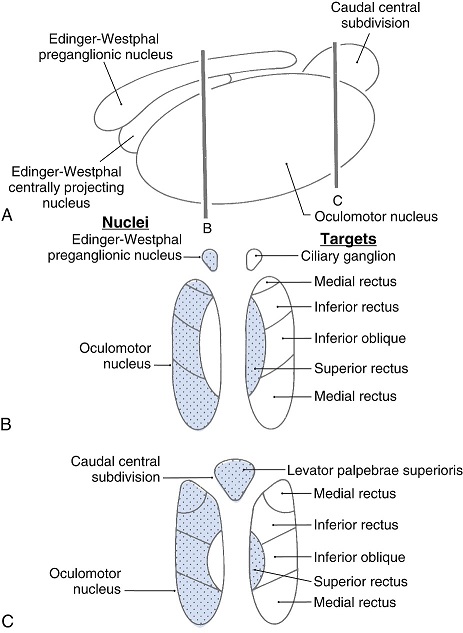 The oculomotor neurons are fast. The figure shows the firing of an abducens motor neuron relative to muscle tension and eye position. Note that the instantaneous firing rate of the motor neuron closely resembles the tension in the eye muscle. However this tension adapts very quickly to eye position, as shown in the top trace. The majority of the saccade-related motor neurons in the adbucens nucleus supply fast twitchy muscle fibers in the lateral rectus muscle. There is also another distinct group of abducens motor neurons that are slighly slower and drive multiply innervated muscle fibers that are more closely related to fixation.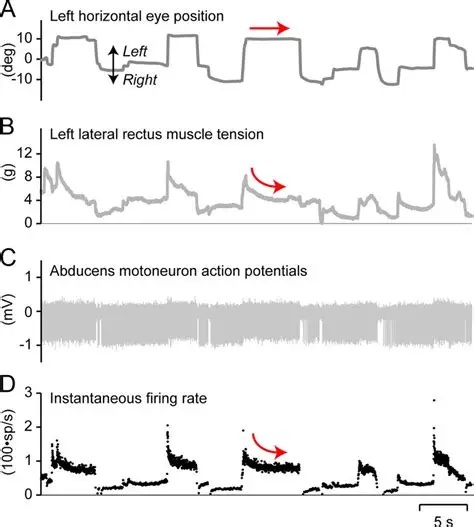 (figure from Lopez-Carrizosa et al 2011) There are four types of neurons in the abducens nucleus, identified by chemical means (Horn et al 2018). Two of them innervate the slow (multiply innervated MIF) and fast (singly innervated SIF) muscle fibers of the lateral rectus muscle. These neurons are both cholinergic. A third type, an interneuron, crosses the midline to connect with the contralateral oculomotor nucleus via the medial longitudinal fasciculus. It is not cholinergic. These interneurons get inputs from the same set of PPRF premotor burst neurons as the abducens motor neurons themselves, including the integrators in the NPH and MVN, and the pause neurons in the NRI (please refer to the glossary for a review of these terms). The abducens interneurons drive the contralateral eye for conjugate eye movements. A fourth group of AChE-positive (but ChAT-negative) neurons, located rostrally and near the midline, connects with the flocculus and ventral paraflocculus of the cerebellum via the paramedian tract. These PMT neurons contribute to gaze holding. The SIF neurons are clustered in the middle of the adbucens nucleus, and the MIF neurons form a shell around them. The PMT neurons form a rostral cap on the entire structure.The palisade endings on the eye muscles are an anomaly. Early research in cats and monkeys indicated they're cholinergic and originate from the motor neurons themselves, but later research seems to point to a small separate cell group within the motor neuron population. The palisade endings are anomalous because the fast twitchy SIFs in monkeys exhibit the classical en plaque endings near their midpoints, and the MIFs show small en grappe endings that are distributed along the whole length of the muscle fiber (Spencer and Porter 2006). The palisade endings may serve a motor related role, but it is equally likely they somehow deliver proprioceptive information back into the oculomotor system. The figures shows some palisade endings on a lateral rectus muscle.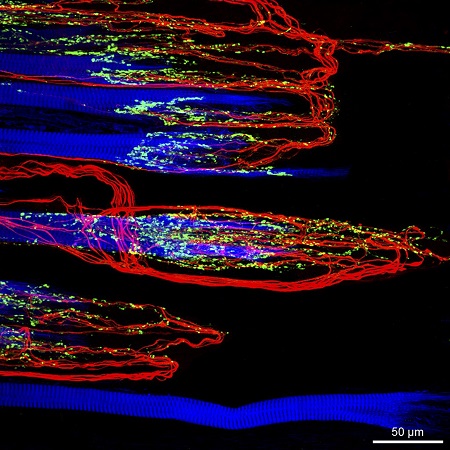 (figure from Streicher et al 2022) In addition to the oculomotor neurons themselves, there are motor neurons responsible for contracting the pupil, accommodating the lens, and raising the eyelid. Pupil constriction is a parasympathetic response driven from the nucleus of Edinger-Westphal. There is a light-based reflex originating in the retina, fibers from retinal ganglion cells travel through the optic chiasm and optic tract before entering the pretectal nucleus (specifically the subdivision called the pretectal olivary nucleus), and from there the neurons connect with Edinger-Westphal, which sends preganglionic fibers to the ciliary ganglion, which in turn sends postganglionic fibers to the sphincter pupillae (a circular muscle in the iris), causing the pupil to constrict (Akova et al 2025). The pretectal nuclei drive the nuclei of Edinger-Westphal bilaterally, so a light stimulus in one eye will cause both pupils to constrict. These pathways are shown in the figures.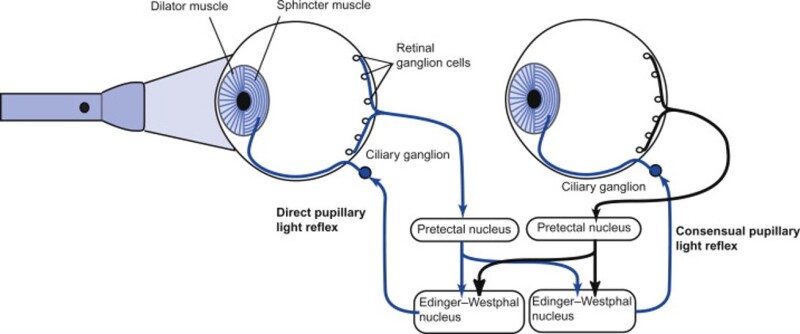 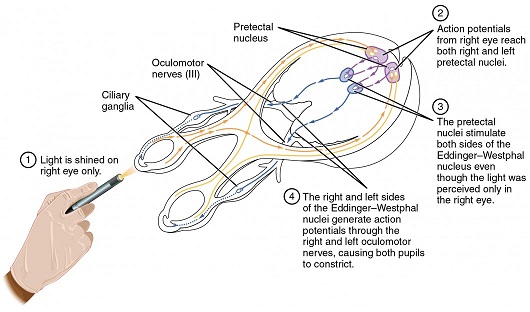 The lens accommodation pathway drives the ciliary muscles. These muscles contract for near objects (causing the lens to become thicker and more spherical, increasing its refractive power), and relax for distant objects. Lens accommodation is a parasympathetic response. There is an accommodation reflex that includes the pupil and medial rectus muscles as well as the ciliary muscle. These three components are called the "near triad", as accommodation usually includes ocular convergence (an inward movement of both eyes), and miosis (pupil constriction to increase the depth of focus). Since lens accommodation is related to depth perception, there are control signals issued from cerebral area 19 (V3) that travel to the pretectal area and then to the nucleus of Edinger-Westphal, which sends fibers through oculomotor nerve into the ciliary ganglion, which in turn feed the ciliary muscles of the eye. The midbrain center for the near reflex (in the pretectal area) is more ventral than that for the light reflex driving pupil constriction. It is a separate area that organizes the near triad. Within the pretectal area, it it closer to a region that involves the nucleus of the optic tract and the posterior pretectal nuclei. The nucleus of the optic tract receives direct retinal projections and is also involved in optokinetic nystagmus, short-latency ocular following, smooth pursuit eye movements, and gain adaptation in the vestibulo-ocular reflex (Gamlin 2006). Burst NeuronsExcitatory burst neurons (EBN's) drive both the oculomotor neurons and the oculomotor integrator. This pattern is shown in the figure below. The burst neurons become active immediately (about 12 msec) before an ipsiversive saccade, and their firing rate is predictive of eye velocity. The burst neurons fire only during rapid eye movements (saccades) and do not activate during fixation, pursuit, or vergence movements. (There is debatably a separate set of burst neurons related to vergence in humans, although that is still controversial).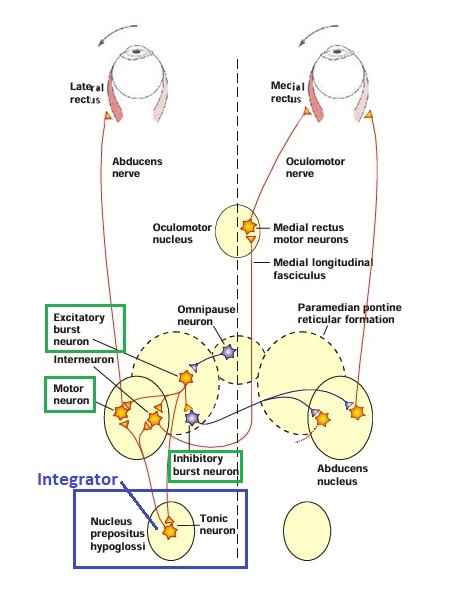 The EBN's for horizontal saccades are located in the paramedian pontine reticular formation (PPRF). Those for vertical saccades are located in the interstitial nucleus of the medial longitudinal fasciculus (riMLF). The excitatory burst neurons generally issue collaterals that cross the midline and directly excite the opponent muscle of the contralateral eye. For example when the right eye wants to move to the right, the left eye must move to the right also and therefore a contraction of the abducens muscle on the right must be paired with a contraction of the oculomotor muscle on the left. Oculomotor IntegratorThe oculomotor neurons are tonically active. The eye muscles must maintain a small amount of force to properly balance the eyes. The baseline firing rate of the motor neurons is related to eye position. To maintain a firing rate related to eye position even when there are no incoming eye movement commands, an integrator is necessary. The integrator sets the new baseline firing rate according to the final eye position. To accomplish this, the integrator naturally integrates velocities, essentially like a path integral, where the amount of travel relates to the distance of the target from the current eye position. When the eyes return to their resting positions, the oculomotor integrator will need to be reset, and there are several ways this can be done. 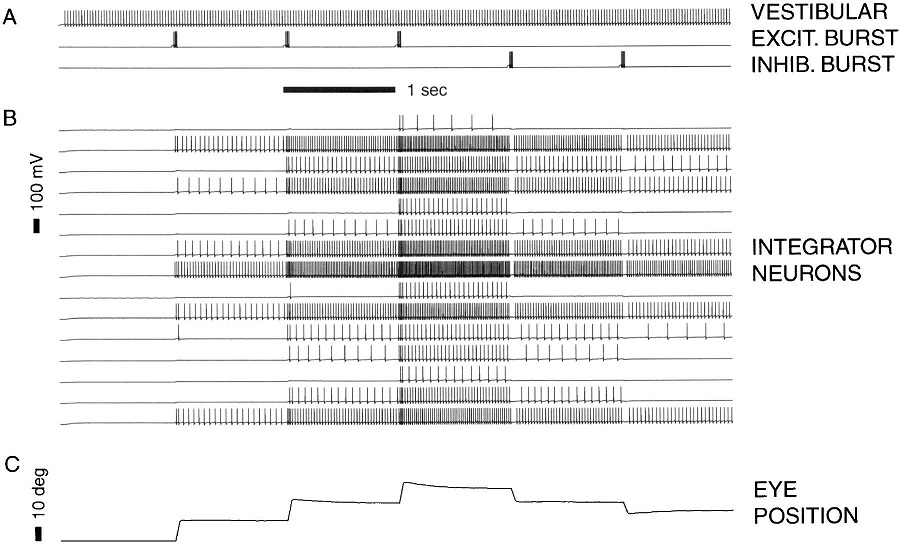 (figure from Seung et al 2000) The step response of the integrator is thought to be accomplished through recurrent excitatory connections within the integrator. This architecture is shown in the figure. It has been successfully modeled by Seung et al using conductance based neurons. (While the figure is reminiscent of the Hopfield network shown earlier, the oculomotor integrator is most definitely not a thermodynamic network and should not be confused with a Hopfield machine!). In this diagram the gain of the integrator is determined by the strengths of the recurrent synaptic connections. These connections can adapt "such that" the gain is correct, provided there is a closed loop error signal from the visual image (target) back into the oculomotor system. Such a signal can be derived monosynaptically from the connection between the retina and the superior colliculus, and polysynaptically in many ways by connections from the visual cortex into the oculomotor system.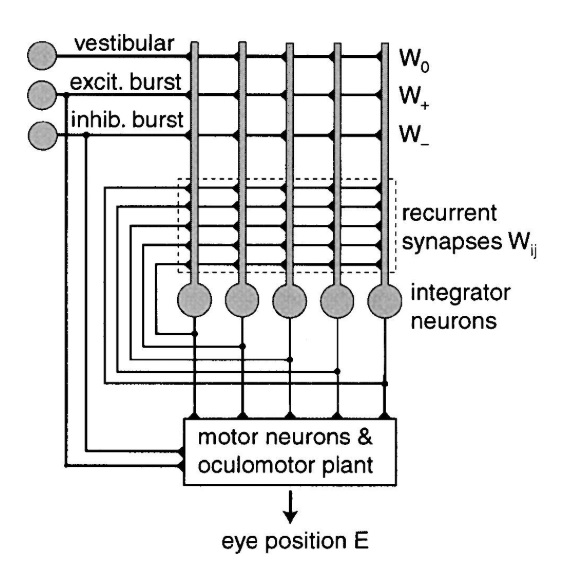 (figure from Seung et al 2000) Single neurons within the integrator display mostly linear relationships with eye position, and they have a threshold position below which they do not activate. This behavior is shown in the figure. (In this figure showing horizontal movements, B are the burst neurons and OI are the oculomotor integrators).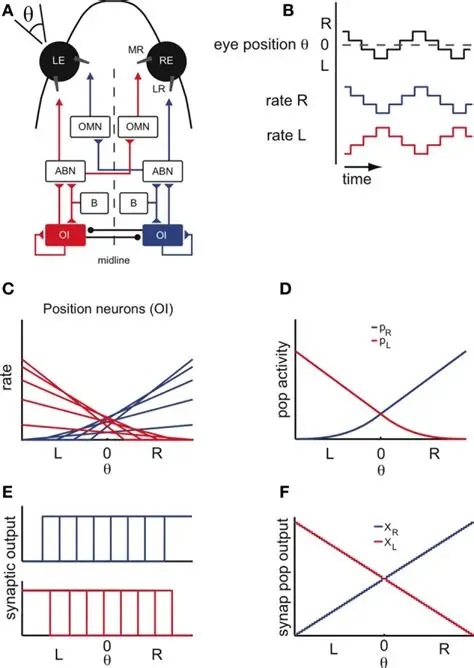 (figure from Goncalves et al 2014) This behavior is reproduced quite nicely in the model by Seung et al. One of the interesting things about this model is the instantiation of the "encoders" described in Nengo's Neural Engineering Framework. In this case, the interpretation of encoders dovetails quite nicely with the variability found in real biological systems.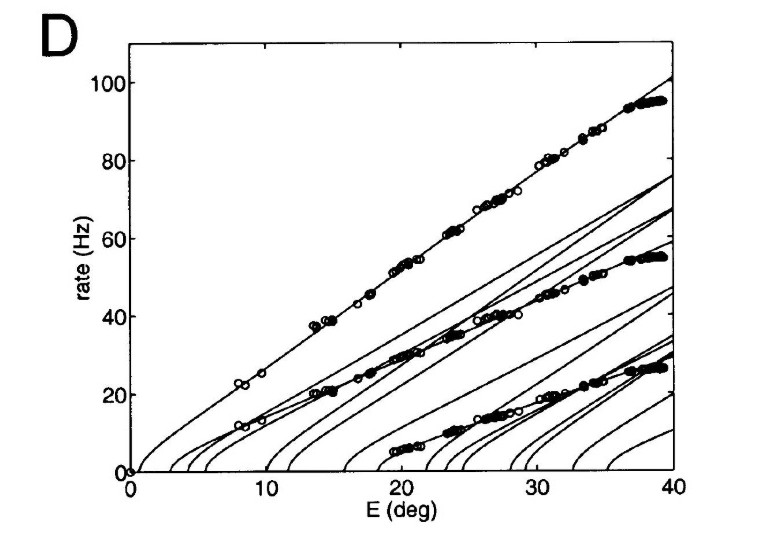 (figure from Seung et al 2000) Below is a control systems view of the integrator circuitry. In this figure, OMN are the oculomotor neurons, NI is the integrator, and MLF is the direct synaptic connection from the burst neurons to the motor neurons via the medial longitudinal fasciculus.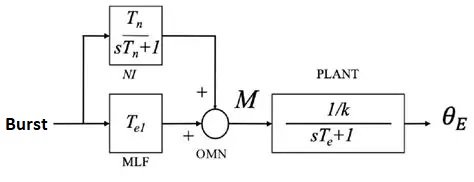 Pause NeuronsIn addition to the burst neurons, there are pause neurons (sometimes called "omnipause" neurons because they inhibit all eye movements bilaterally). The pause neurons are located in the nucleus raphe interpositus. They are inhibitory neurons that use glycine as a neurotransmitter, monosynaptically inhibiting both excitatory and inhibitory burst neurons. The pause neurons fire tonically during fixation, at rates in the 100-200 Hz range, but stop just before a saccade. When the pause neurons become active, they inhibit the burst neurons, so eye movements stop and the eyes stay where they are. Thus the pause neurons typically become active at the end of an eye movement, and remain active during periods of fixation. However note that the pause neurons do not directly inhibit the ocular motor neurons, because the latter need to stay tonically active while the eyes are fixated. Instead, they inhibit the burst neurons that send inputs to the motor neurons. This circuitry is shown in the figure, indicating the monosynaptic inputs from the superior colliculus to the pause neurons, which then inhibit the burst neurons. The figure also shows the timing of neural activity in each of the neurons.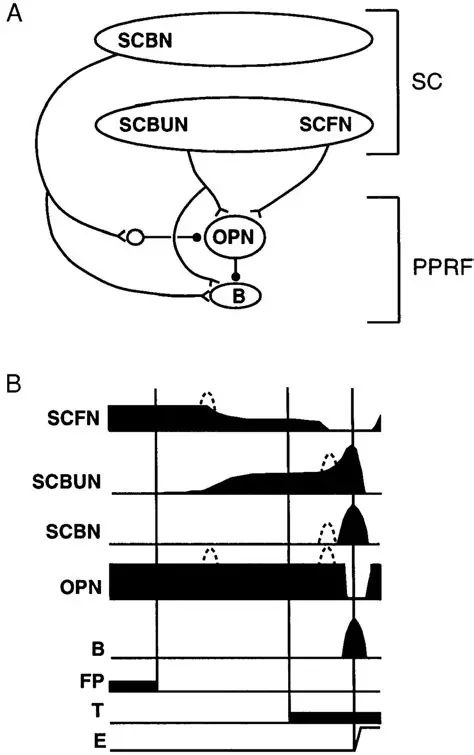 In the figure above, the burst and pause neurons are shown being driven by the targeting system in the superior colliculus (by both bursting neurons and fixation neurons). We will discuss the superior colliculus on the next page. For the drawing, be aware that in the SC, neurons closer to the fovea tend to hold gaze while those in the periphery tend to initiate saccades. Thus the central neurons would excite the pause neurons and inhibit the EBN's, while the peripheral neurons would excite the EBN's and inhibit the pause cells. 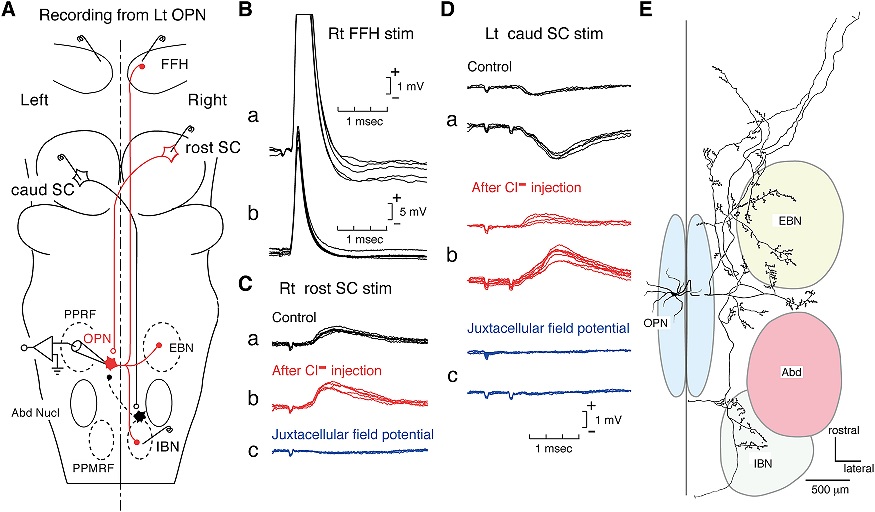 (figure from Takahashi et al 2022) However as shown above, there is also a direct monosynaptic pathway from the superior colliculus to the IBN's, and the IBN's in turn inhibit the pause neurons. The pause neurons and the IBN's are mutually inhibitory. The mutual inhibition between IBN's and pause neurons can play an important role in the decision to initiate a saccade.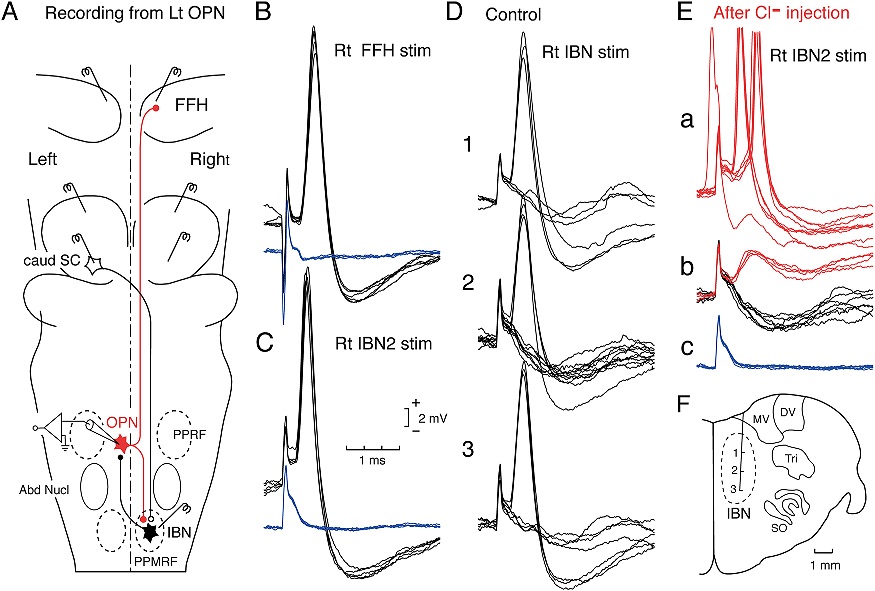 (figure from Takahashi et al 2022) The oculomotor bursting system in the brainstem is potentially unstable, when it's not being inhibited by the pause neurons. When the pause neurons are temporarily disabled, oscillations can be induced in both the bursting neurons and in the resulting eye positions (Ramat et al 2005). The stability of the eyes during fixation requires the pause neurons to be turned on, and it is further refined by input from the cerebellum. The cerebellum performs multiple functions related to eye movements, not only is it responsible for big portions of the vestibular input and the information related to smooth pursuit, but it also fine tunes the eye positions during saccades and keeps the eyes on target.Long Lead Burst NeuronsLong lead burst neurons are believed to live "between" the superior colliculus and the EBN's, synaptically. They likely feed the EBN's in both the horizontal and vertical systems (in the PPRF and riMLF). They have a peculiar pattern of activity, beginning with a low frequency prelude starting up to 150 msec before a saccade, and eventually transitioning into a higher frequency burst. There are likely several classes of such neurons (Walton and Freedman 2013). Target SelectionTarget selection is organized in the superior colliculus, under the influence of inputs from the cerebral cortex, involving primarily the frontal eye fields and the intraparietal eye fields (both part of the dorsal attention network). The superior colliculus brings together multiple cerebral systems for the control of the brainstem oculomotor circuits. The superior colliculus is the subject of the next page.Next: Superior Colliculus |