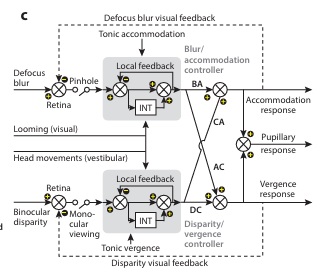
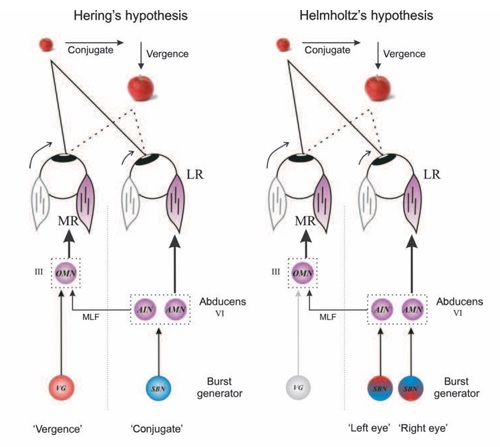
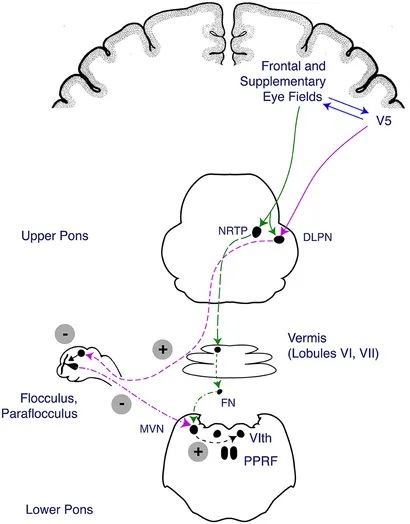
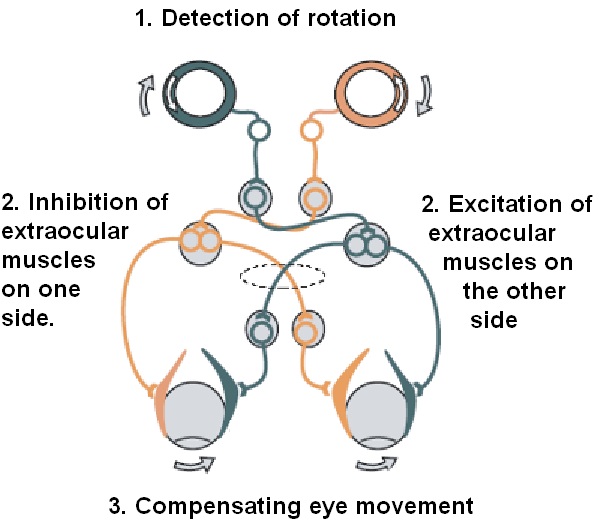
Overview of Oculomotor SystemIn this section we'll review human eye movements and outline the behavior of the eyes for each movement type, along with providing some context for the subsequent sections where we'll discuss targeting and attention. 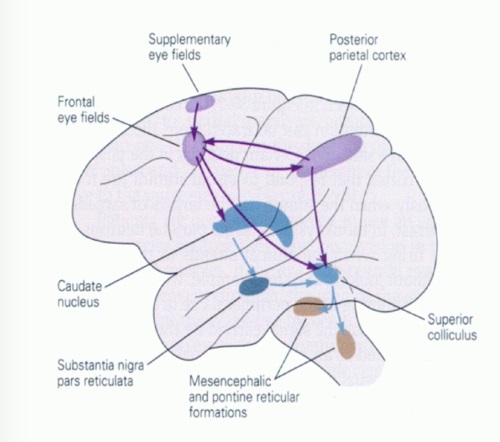 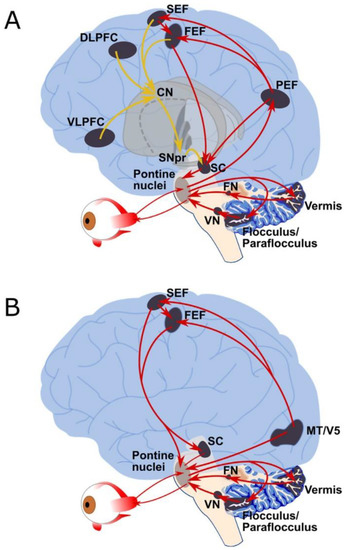 Here is the corroboration of these pathways from the standpoint of functional MRI. The images start at the top left in the brainstem, and proceed to the rostral surface of the brain. On the top left you can see the abducens nucleus, one of the most important oculomotor nuclei in the brainstem that directly drives the lateral rectus muscle. On the top right is the superior colliculus, an important structure for target selection. On the bottom row are the parietal and frontal eye fields, and the supplementary eye fields in the premotor area. The cingulate eye field is shown in light blue in the middle row on the right.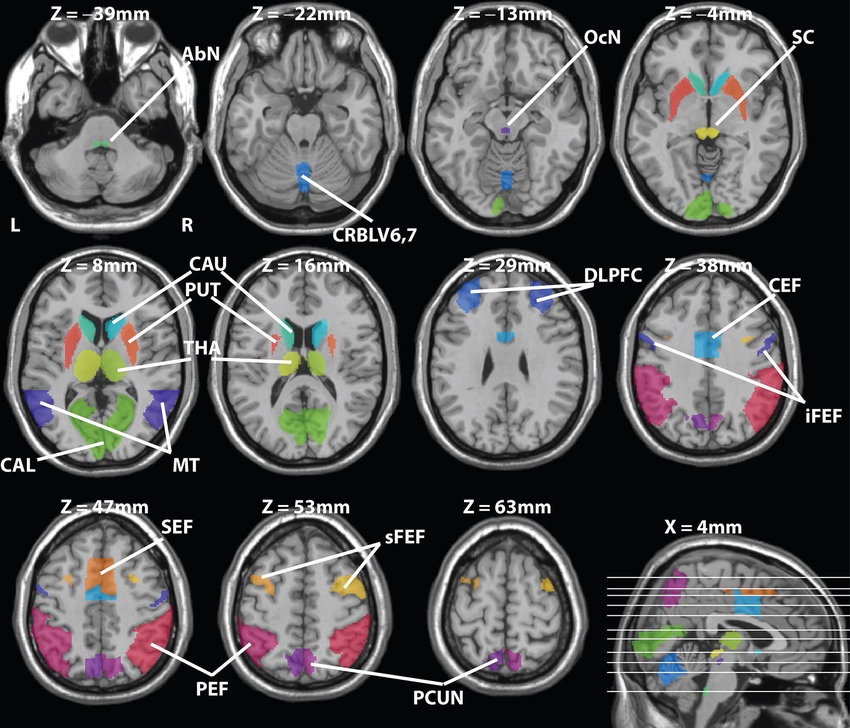 To understand this architecture, we'll work bottom up. We'll start with the eyes and the eye muscles, and work our way up to the cerebral cortex. The organization of the oculomotor system is most logical when understood this way.Eyes and Eye MusclesThere are six main muscles that move the eye in its socket, and several others related to the eyelid, pupil, and lens. Eye movements along the horizontal and vertical axes are organized by the rectus muscles, which are arranged into push-pull pairs. Each muscle maintains a low level of tonic activity even during fixation, to keep the eye stable and avoid drift. The figure below shows the organization of the muscles in the right and left eyes. 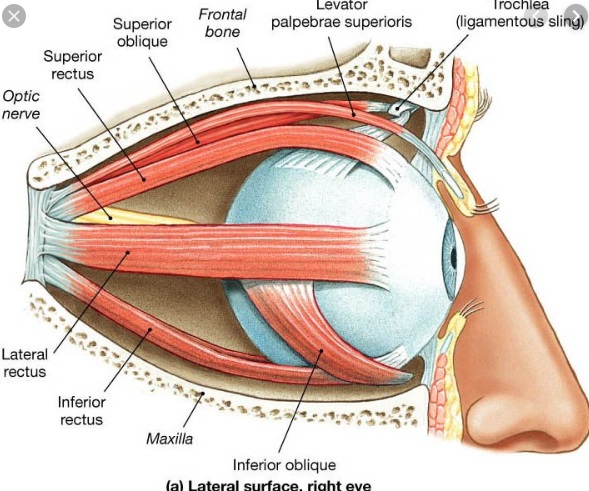 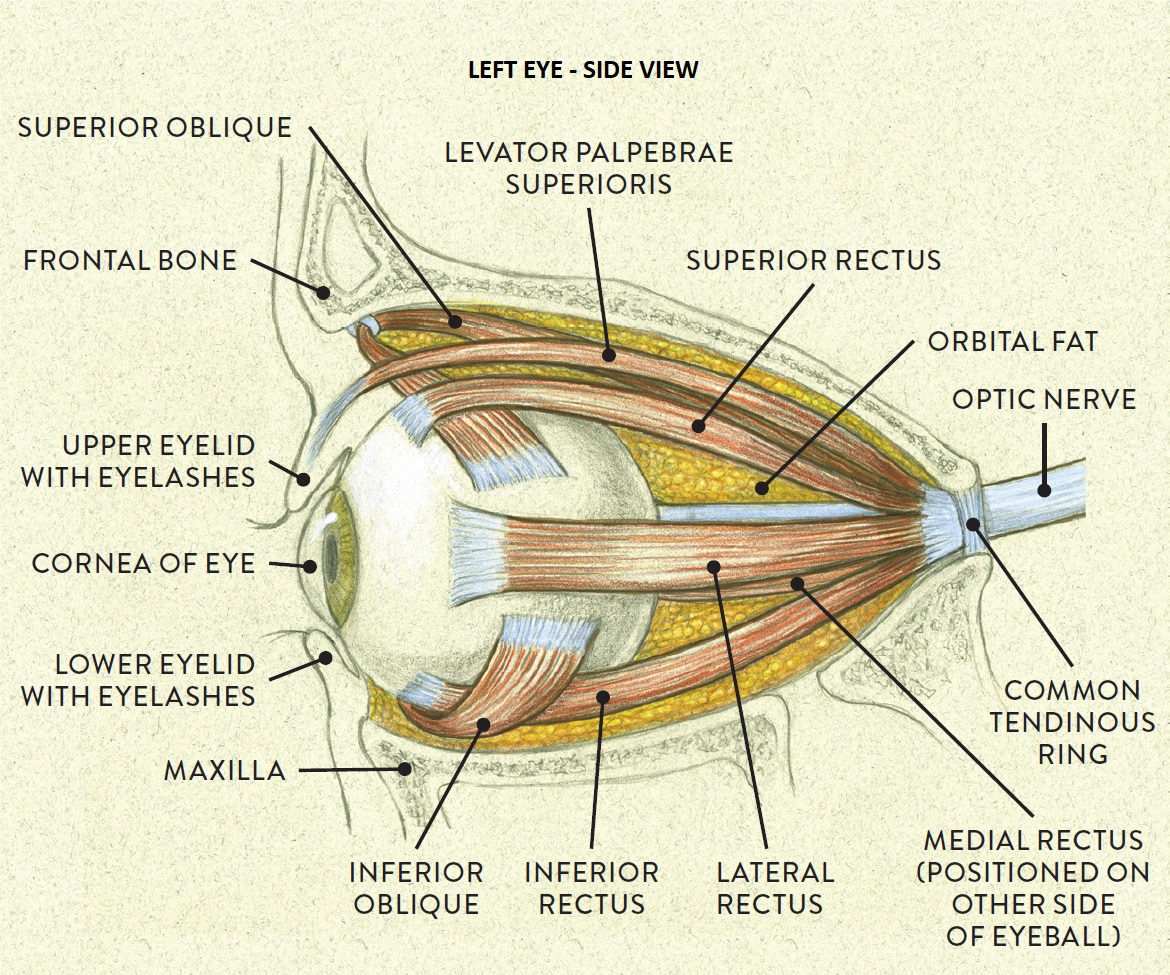 The eyes can move together, conjugate from side to side, or they can move disjunctively, for example when focusing in depth. These systems are mostly separate in the brain, for example there is a separate system for vergence (disjunctive) movements. However they come together at the level of the oculomotor neurons. As you can see from the figures at the top of the page, the circuitry of the oculomotor system is somewhat complex, and the insertions of the eye muscles are not necessarily conveniently aligned along Cartesian axes. However the overall architecture can be organized functionally, into subsystems and pathways that make sense. 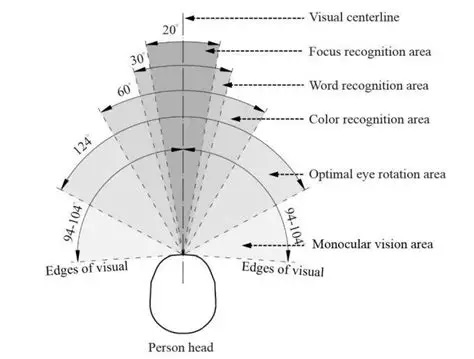 The range of eye movements is approximately aligned with the "immediate field of view", as shown in the figure. Targets outside of this range tend to evoke more complicated orienting movements involving the head and body. Some of these movements are organized by the superior colliculus, and transmitted through descending fibers that target the cervical and spinal motor pathways. There are other pathways through the cerebellum, involved with gaze maintenance and the precision of fixation. These systems are complex and interact with each other at many levels. For example the vestibulo-ocular reflex has to be compensated during tracking movements that involve the head. Much of what we know about human eye movements comes from clinical conditions where either the range, velocity, or behavior of the eyes (or all of these) is altered.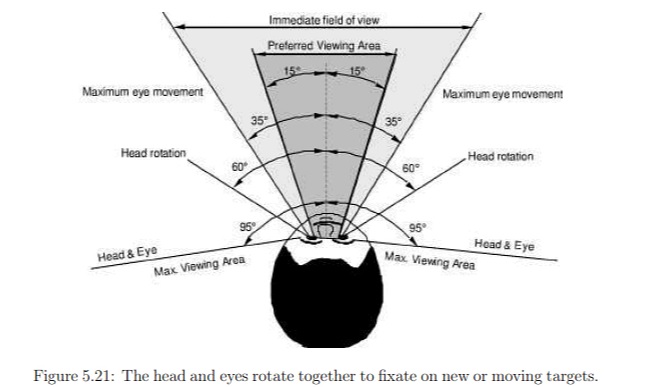 Overview of Eye Muscle ControlAnatomically, much of the eye movement circuitry is organized at the level of the midbrain, in the pons and neighboring areas, close to the vestibular systems and close to the cerebellum. The pontine reticular formation is an intricate tangle of tiny nuclei and fibers going in all directions. It has taken many years of study, and the invention of some clever technology, to map this area of the brain. The basic circuitry around the abducens nucleus is shown in the figure. You can see the pathway that cross the midline and activates the oculomotor nucleus on the contralateral side, which contracts the contralateral medial rectus muscle and keeps the eyes in alignment. A lesion or tumor at position 4 will affect the movement of both eyes in the contralateral direction, whereas cutting the abducens nerve at position 1 affects only one eye - however the brain tries to compensate for oculomotor deficits, and the time course of adaptation is an important clinical indicator.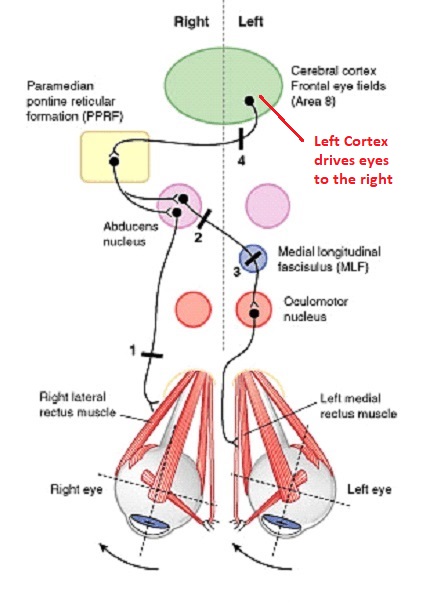 The eyes have "focus", both horizontally and vertically, and in depth. The horizontal and vertical foci are often conceived in terms of "gaze centers". When organized this way, the horizontal gaze center is in the paramedian pontine reticular formation (PPRF), and the vertical gaze center is in the rostral interstitial nucleus of the medial longitudinal fasciculus (riMLF). As you can see in the figure, horizontal saccades are primarily driven by the lateral rectus muscle on the abducting side, and its accompanying neural circuitry. The PPRF contains bursting neurons that drive the motor neurons in the abducens nucleus during saccades. Both of these areas get direct input from the cerebral cortex and the cerebellum. As a general rule, the firing rate of abducens motor neurons is directly related to eye position, whereas the bursting neurons convey velocity signals, that need to be integrated to translate them into eye positions.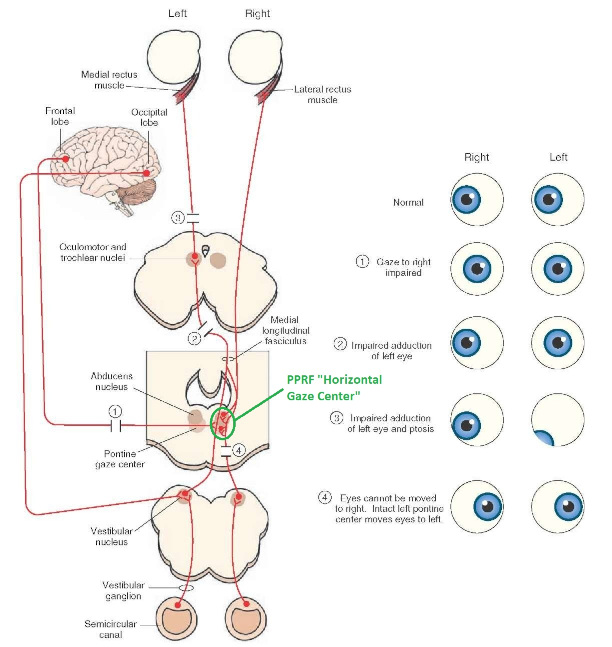 Vertical saccades use a separate set of pathways that control the superior and inferior rectus muscles via the oculomotor nucleus, whose axons travel through cranial nerve 3. Because of the way the eyes are arranged in the orbit, vertical eye movements also generate small amounts of torsion, and to correct this the superior and inferior oblique muscles get involved too. The superior oblique has its own pathway through the trochlear nucleus and the trochlear nerve (cranial nerve 4), whereas the inferior oblique uses the oculomotor nucleus and CN-3. When the eyes are at rest, the tension on each muscle is minimal and so is the firing of the motor neurons that drive the muscles. Whenever the eyes deviate in any direction, they tend to eventually slip back into their resting positions. To maintain them in an eccentric position, the motor neurons have to keep firing, and that is part of what "gaze" is, it's a careful adjustment of motor neuron firing rate to keep the eyes on target. Without continuing tension, the retinal image will slip, creating an error signal in the targeting system.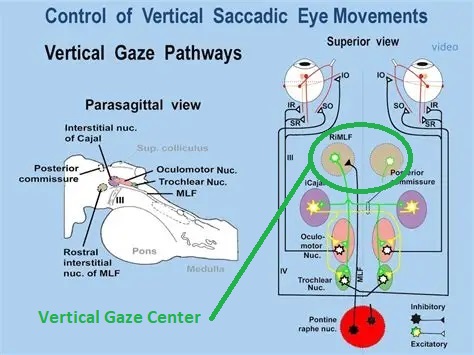 When the eyes are focused together on a target (moving or not), the angle between the eyes is calculated in relation to depth perception. Movements in depth are vergence eye movements, and they are regulated by a separate system in the brain. In general, each axis of motion is regulated by separate systems. The vergence system issues "corrections" that enter at the level of the oculomotor nuclei. These affect the oculomotor nucleus specifically, and not the abducens. Thus, human vergence movements often involve both eyes moving together towards the midline. The circuitry involves the supra-oculomotor area including the nucleus of the posterior commissure (Friedrich et al 2026). In humans, vergence movements have fast and slow components (Cullen and Van Horn 2011), they are guided by the cerebellum.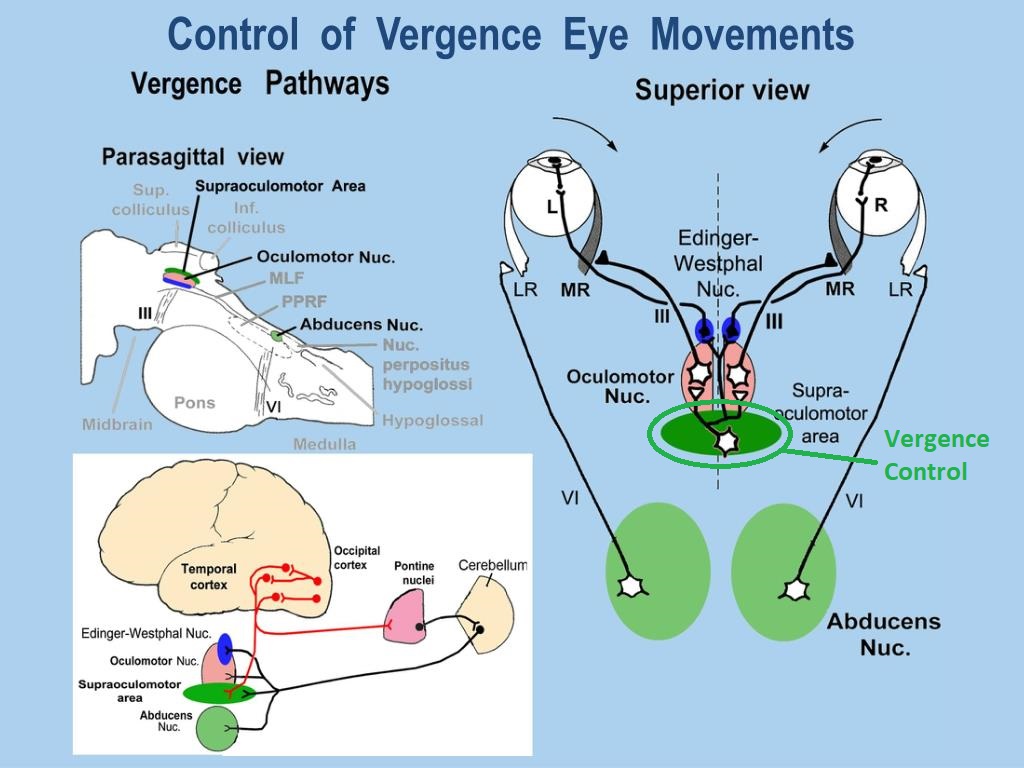 In addition to these three major axes of motion (horizontal, vertical, and vergence), there is a lot of reflex-related circuitry pertaining to the relationship between the eyes and head, and the eyes, head, and visual targets. There are reflexes related to pupil constriction, lens accommodation, and eye blinks, each of which have their own set of pathways. Higher level reflexes like the looming reflex involve the same targeting system that orients the organism in relation to the visual field. Each of the processing centers and connection pathways has a specific function, and often the subsystems end up working together to accomplish sophisticated behaviors. As a general rule the cerebellum is reciprocally connected at every level of the peripheral oculomotor system, whereas the cerebral cortex mainly provides input (its return information comes in indirectly, through the thalamus, both from visual information derived from the retina and from targeting information supplied by the superior colliculus to the pulvinar nucleus).Horizontal SaccadesHorizontal eye movements involve the lateral and medial rectus muscles. The primary driver for the saccade is the lateral rectus muscle on the ipsilateral side (the one moving away from the midline). The oculomotor nucleus plays a somewhat secondary role in horizontal saccades, as it is driven contralaterally from the abducens motoneurons. This arrangement is shown in the figure. Both abducens and oculomotor nucleus neurons are eventually driven from the superior colliculus, but along the way the signals are processed by small brainstem nuclei that translate the desired eye position into actual muscle contractions. In humans there are about 6500 neurons in the abducens nucleus. 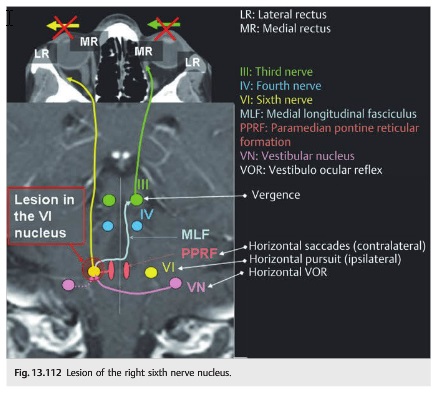 Abducens motor neurons are driven by excitatory bursting neurons (EBNs) in the ipsilateral paramedian pontine reticular formation (PPRF), and are inhibited by pause neurons in the nucleus raphe interpositus (NRI). There are several other direct inputs, including most importantly the medial vestibular nuclei (for the vestibulo-ocular reflex), the nucleus prepositus hypoglossi (related to the neural integrator for horizontal eye movements), the interstitial nucleus of Cajal (related to vertical and torsional eye movements), the supraoculomotor area (related to vergence movements), and the pretectal area surrounding the superior colliculus (Buttner-Ennever 2006). The motor neurons in the adbucens nucleus are the final common pathway for all of these eye movement signals, and in many cases other types of eye movements (like vergence movements) have to continue during saccades, so they are additive at this level. The burst neurons in the PPRF convey signals specifically related to saccades. They burst just before the onset of a saccade. Saccades are "released" by a removal of inhibition from the pause neurons in the NRI. The burst and pause neurons work together to determine the exact timing of a saccade. The schematic below shows excitatory bursting neurons (EBNs) from the PPRF driving the abducens motor neurons. It also shows two types of oculomotor integrators, one in the nucleus prepositus hypoglossi related to saccades, and another in the medial vestibular nucleus related to the VOR and target pursuit during head movements.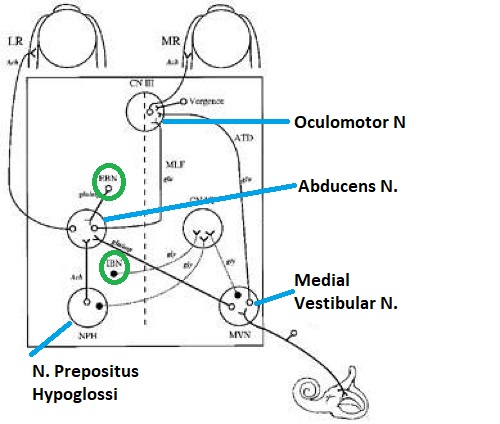 There are four sets of outputs from the abducens nucleus, from four different kinds of neurons. Two of them drive the twitchy and non-twitchy lateral rectus muscle fibers directly, the third crosses the midline to drive the contralateral medial rectus muscle, and the fourth sends axons into the paramedian tract to connect with the flocculus of the cerebellum. Each of these neurons receives a different kind of input. The twitchy motorneurons (related to saccades) get most of their inputs from the EBN's in the PPRF, while the non-twitchy motorneurons (related to gaze holding) are driven primarily from the NPH (Horn et al 2018). Incoming fibers from the medial vestibular nucleus (transmitting velocity signals for the VOR) cross the midline and excite both the contralateral twitch neurons and the contralateral internuclear neurons. 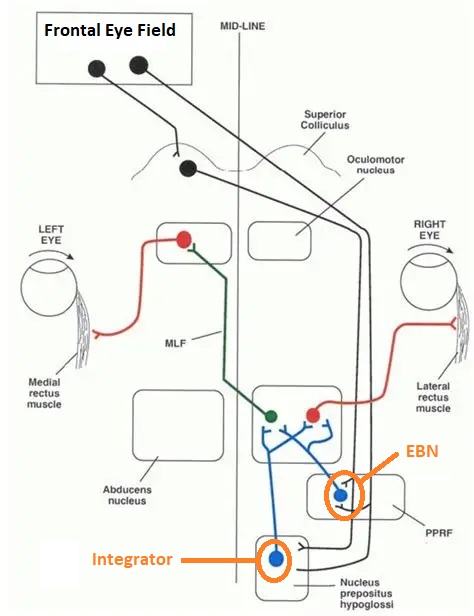 Saccades engage both eyes. The movements are "conjugate", and so is the circuitry. There are excitatory and inhibitory burst neurons driving saccades. The excitatory burst neurons excite the abducens motor neurons on the same side (ipsilateral), and the oculomotor neurons driving the medial rectus muscle on the opposite side (contralaterally). At the same time, they connect with inhibitory burst neurons on the opposite side, which inhibit the abducens motor neurons contralaterally, and the medial rectus ipsilaterally. These connections are shown in the diagram below. The burst neurons are located in the paramedian pontine reticular formation (PPRF), and the pause neurons are nearby in the nucleus raphe interpositus (NRI). EBNs from the PPRF project to the contralateral IBN's located in the area of the nucleus paragigantocellularis. The neurons labeled INT are the non-cholinergic abducens interneurons that cross the midline to feed the contralateral medial rectus motor neurons. Not shown are the connections with the cerebellum.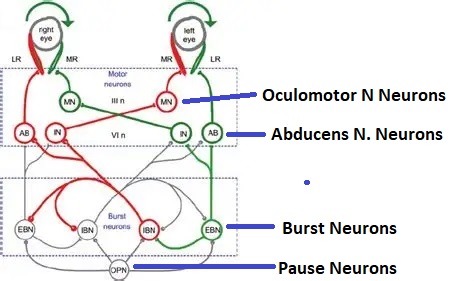 The firing pattern of an abducens motor neuron and an excitatory burst neuron relative to a saccade is shown in the figure below. Note that the burst occurs slightly before the saccade, and that the abducens motoneuron retains a new tonic firing level after the saccade. In this case the number of spikes in the EBN burst is linearly related to saccade amplitude (although the abducens motor neurons shows interesting step-like behavior)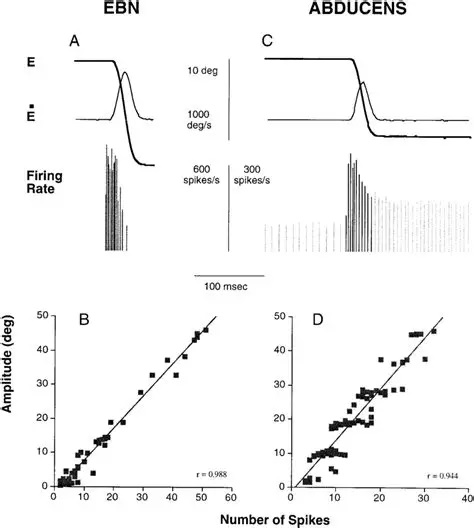 Between the burst neurons and the oculomotor neurons is a neural integrator (appropriately labeled in the previous two diagrams), that performs two essential functions. First, it converts the spike train representing a saccade into an eye position, which is a level of muscle contraction. And second, it maintains the eye position in the face of noise and small changes in muscle activity. In humans this integrator is located at least in part in the nucleus prepositus hypoglossi, which was shown in an earlier cross section of the pons (above). The NPH works in conjunction with the medial vestibular nucleus, which receives head velocity signals from the semicircular canals. The oculomotor integrator performs a crucial function for both saccades and tracking, in that it translates the velocity information into eye position. The integrator is still controversial. There are many ways to accomplish integration in neural networks, and in humans the wiring is such that there are likely to be multiple integrators that interact with each other. For example the horizontal and vertical integrators are not cleanly separated, horizontal saccades typically produce tiny vertical and torsional movements as well, and vertical eye movements often result in immediate horizontal corrections. Integrators can be either feed-forward or recurrent, and both varieties have been proposed in various combinations to account for human eye movements. The figures show these two arrangements, in the first version there's a feed-forward integration pathway that's supposed to resemble the anatomy in the nucleus prepositus hypoglossi (NPH), and in the second version there's a feedback pathway from the abducens through the cerebellum and back into the burst neurons. Both pathways may exist in a human brain.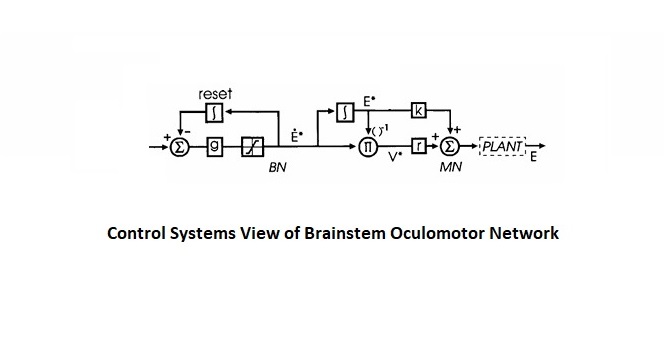  (figure from Kheradmand and Zee 2011) There is something missing in the above diagrams. There are muscle spindles in the eye muscles, however they're controversial too, and they look a little different than they do in ordinary skeletal muscles. In humans and primates, there are specialized structures called "palisade endings" that are thought to provide proprioceptive input, even though they seem to originate from the motor neurons themselves. The palisade endings are about halfway between ordinary muscle spindles and Golgi tendon organs. They connect mostly with the smoother slower non-twitchy eye muscles. There is some evidence that the cell bodies giving rise to palisade endings are distinct from the motor neurons, the motor neurons are large and cholinergic whereas the PE cells are round and calretinin positive (Leinbacher 2012). A color picture of the palisade endings is shown on the next page. In the figure below, SIF is a singly innervated muscle fiber of the fast twitchy variety, and MIF is a slower multiply innervated muscle fiber. As described on the next page, the MIF motor neurons terminate in multiple en grappe endings along the entire length of the muscle fiber, while the SIF has a more classic synaptic plaque near the middle of the muscle fiber.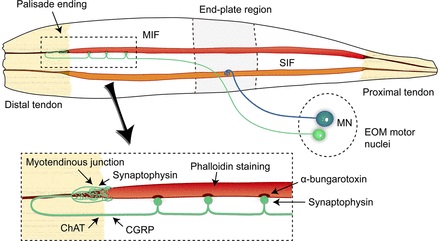 There is some evidence of specificity in the connections from the upstream areas (PPRF, NPH, MVN) into the abducens motoneurons. The faster twitchy neurons get burst input from the PPRF and vestibular nuclei, that generates eye movements. The slower non-twitchy neurons get eye position information from the neural integrator in the NPH. This specificity is currently the subject of active research. Vertical SaccadesThe range of vertical eye motion in humans is slightly smaller than the range of horizontal motion. Vertical eye movements involve the superior and inferior rectus muscles, in cooperation with small adjustments from the oblique muscles. 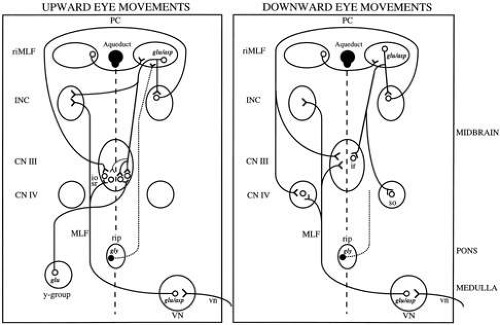 It is important to understand that the separation of horizontal and vertical axes begins all the way out in the semicircular canals. We'll take a moment to review the organization of the vestibular system, because it helps us to better understand the oculomotor design. 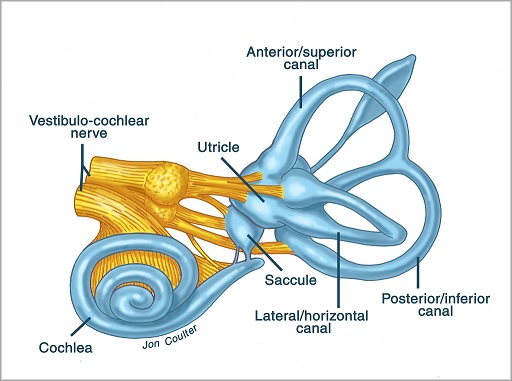 The horizontal semicircular canal is approximately aligned with the horizontal axis of the head and therefore the horizontal axis of the eyes. The superior and inferior canals detect rotation of the head in the sagittal and frontal planes, and they're not "perfectly" vertical, such a motion would engage both of these canals. The primary sensory neurons for the semicircular canals are in Scarpa's ganglion, which is in the internal auditory canal where the superior and inferior divisions of the vestibular nerve converge. These are bipolar neurons, the sensory side (dendrite) travels through the vestibular nerve to connect with the hair cells in the labyrinths, and the motor side (axon) connects with the one of the four subdivisions of the vestibular nucleus.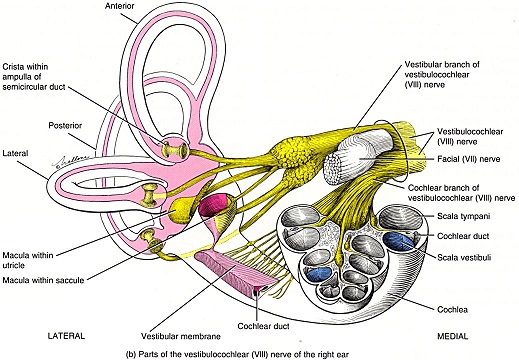 The vestibular nerve (cranial nerve CN-8) has three terminal branches (superior, inferior, and posterior) which innervate the utricle, saccule, and semicircular canals respectively. Moving centrally, they converge in the superior (Scarpa's) and inferior vestibular ganglia. From there they target the vestibular nucleus. There are a few vestibular fibers that target the cerebellum directly, especially the nodulus and uvula of the vermis. These are related to balance and do not primarily affect the oculomotor circuitry.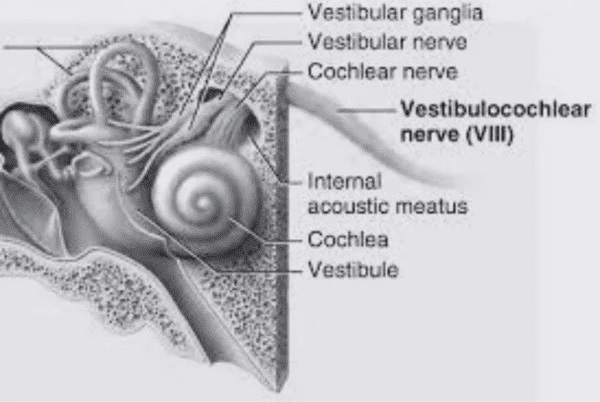 The organization of the vestibular nucleus is shown in the figure. There are four sub-nuclei, called inferior, medial, lateral, and superior. (The lateral subdivision is also called Deiter's nucleus, and the superior subdivision is also called the nucleus of Bechterew, while the inferior subdivision is sometimes referred to as the descending vestibular nucleus or the nucleus of Roller). The main projections of these areas are to the spinal cord (mostly reflexes), to the oculomotor nuclei (for eye movements), to the thalamus (VPi) for perception, and to the cerebellum (for coordination of posture). The medial vestibular nucleus (MVN) is the one that connects with the nucleus prepositus hypoglossi (NPH). It forms an essential part of the oculomotor integrator.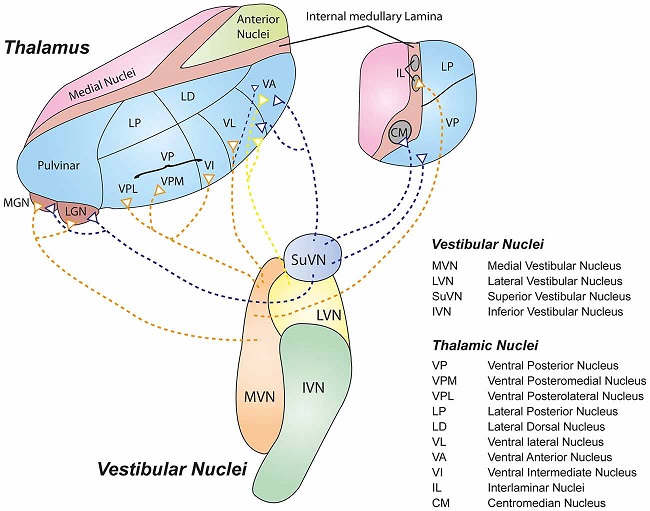 Afferents to the oculomotor system arise from the medial and superior vestibular nuclei. These coordinate eye movements in relation to head movements. The fibers from the superior vestibular nucleus primarily target the thalamus, but they send collaterals into the oculomotor system. The fibers from the medial vestibular nucleus primarily target the oculomotor nuclei. The figure below shows how the fibers from the semicircular canals enter the medial vestibular nucleus, and from there the MVN connects with all of the oculomotor nuclei.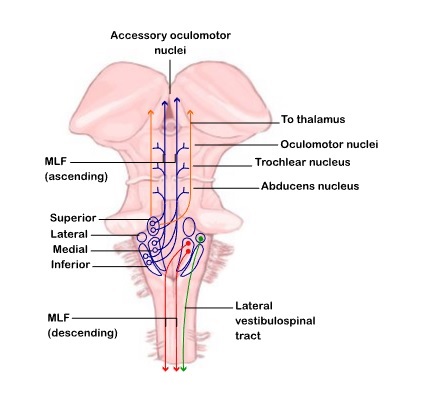
|